Atlas of the Head of a Newborn Wombat (MS363)
Introduction
The common wombat (Vombatus ursinus) is a quadrupedal, burrowing marsupial, mainly crepuscular and nocturnal, found in a variety of habitats (forest, montaine, heathland) in southern and eastern mainland Australia and Tasmania. Adults may be up to 1.0 m in length and weigh up to 35 kg.
Methods
The specimen illustrated here is MS363 (Phascolomys mitchelli) of the Hill collection stored at the Museum für Naturkunde in Berlin. It was a male of 17.5 mm GL (greatest length). The specimen was embedded in paraffin wax and sectioned transversely at an unknown thickness (but possibly 20 µm), before being stained with hematoxylin and eosin. Some of the sections near the edge of the coverslip have been affected by dye leaching and have lost much of their hematoxylin.
Sections at approximately 30 slice intervals were photographed with the aid of a Zeiss Axioplan2 fitted with an AxioCam MRc5 camera. Sections have been depicted in the plates starting with the nostril. All images were calibrated by photographing a scale bar at the same magnification. Images were placed in Adobe Illustrator 2023 and delineated. Developmental regions (i.e. neuroepithelium) destined to give rise to adult structures have been denoted by the adult structure’s name with an asterisk (e.g. Cx* denotes the developmental field of the cerebral isocortex).
An image of the profile of a newborn wombat head was used to construct a line diagram for the finder illustration indicating section position.
Notes on the specimen
General observations
The head of this specimen is very robust and heavy-set (see inset cartoon in each plate depicting the profile of the head) and the cartilage model of the skull base is very thick (see for example the presphenoid - PSph - in plates 4 and 5). This suggests that even a newborn wombat shows adaptations to a life of chewing tough, fibrous vegetation. The developing wombat brain is also very broad (cf. that of the newborn brushtail possum or bandicoot on this website).
Central nervous system
Apart from the broadness noted above, the overall features of the developing brain are similar to that in other newborn diprotodontid marsupials. The telencephalon has a thin pallium (Cx, Cg and Hi), with a thin external preplate (plates 4 to 6) over the dorsal and medial pallium. The lateral cortex (both isocortex and piriform cortex) is better developed, but also thin and immature. The hippocampus (plates 5, 6) is also rudimentary, i.e it has a thick proliferative zone, surmounted by a thin preplate zone, which is mostly fibrous with very few neurons. The subpallial telencephalon has the usual lateral ganglionic eminence (lge in plate 5) and medial ganglionic eminence (mge in plate 5). The striatal subventricular zone has not yet developed and although there are some early-generated striatal and pallidal neurons, it is not possible to distinguish internal structure among those neuronal populations. Only the notional positions of striatal and pallidal components have been labelled.
The hypothalamus is divided into peduncular (hy1), tuberal (hy2), and acroterminal (hyat) proliferative components (plates 5 to 7). The rostral parts of the hypothalamus (plate 5) are very rudimentary and have essentially no postmitotic neurons external to the proliferative zones. In more caudal sections there are more postmitotic neurons in the lateral hypothalamus (plates 6, 7).
The diencephalon has an identifiable prosomeric structure. Prosomeres 1 (pretectum), 2 (dorsal thalamus), and 3 (prethalamus) can be identified as proliferative zones, but postmitotic populations are very small, particularly in the dorsal thalamus.
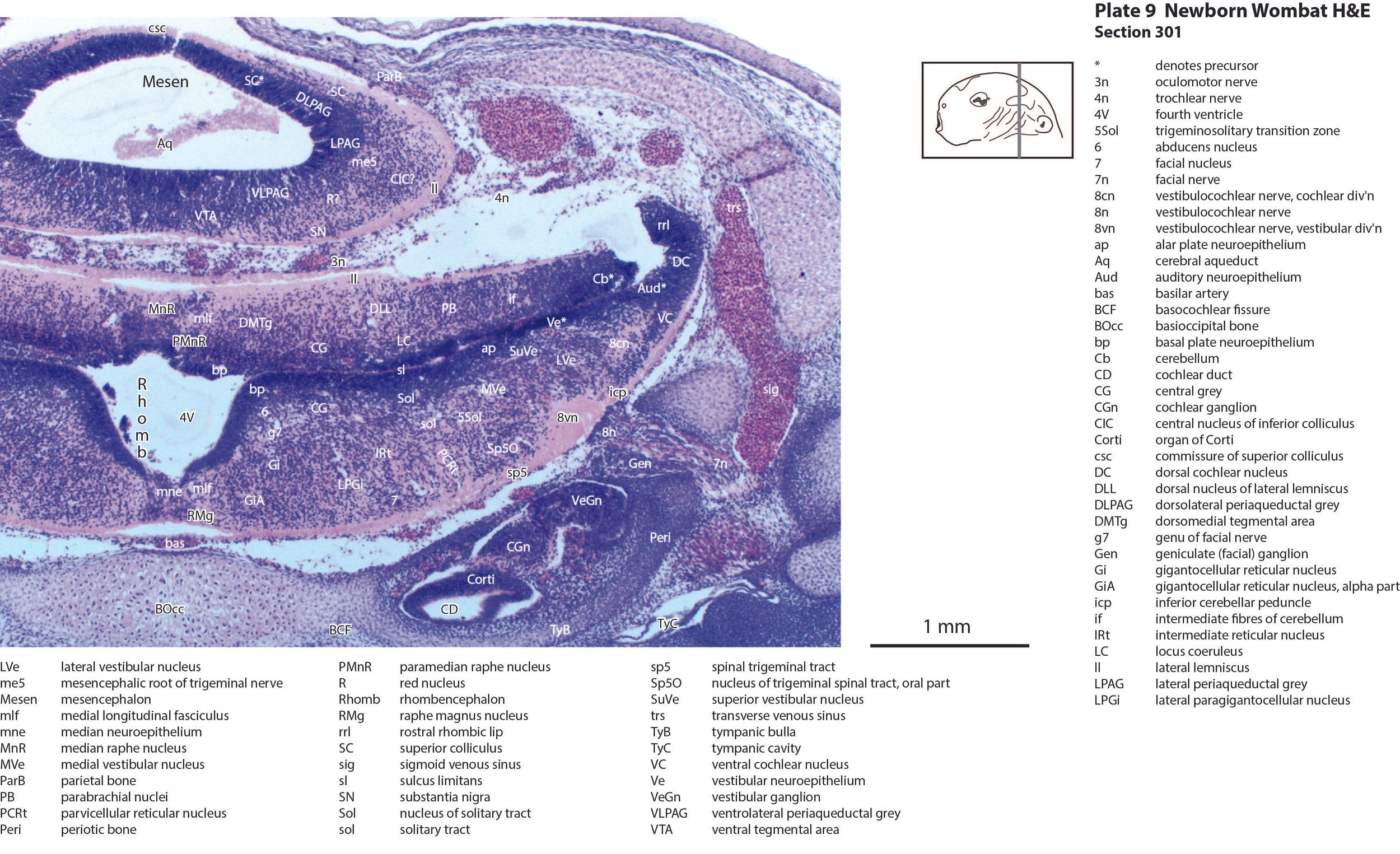
The mesencephalon has a rudimentary superior colliculus (SC in plates 8 to 12) and inferior colliculus (IC in plates 10 to 12), where most of the neuraxis wall is made up of the proliferative neuroepithelium, and very few postmitotic neurons. The mesencephalic tegmentum is also undeveloped. Only the notional position of major nuclei can be marked, because differentiation of nuclei and fibre tracts is poor.
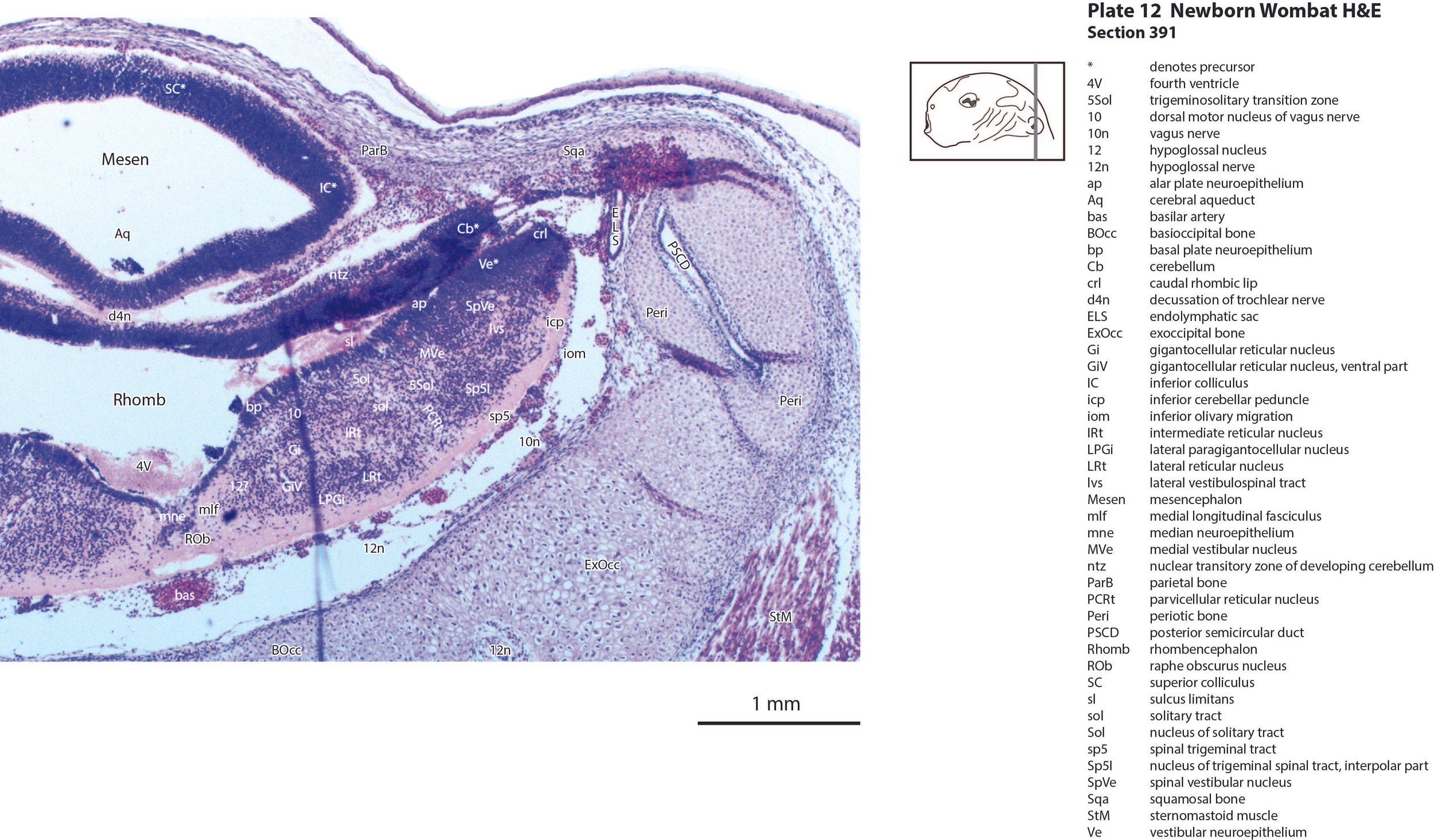
The cerebellum (plates 10 to 13) is a lid extending over the fourth ventricle and rimmed by the rostral rhombic lip. No cortical differentiation and only a nuclear transitory zone (ntz in plates 10 to 12) is evident.
The brainstem of this newborn wombat is at a similar level of differentiation to the newborn brushtail possum. The trigeminal sensory nuclear complex (Pr5, Sp5O, Sp5I; plates 8 to 13; Sp5C not shown) is easily distinguished and the medial magnocellular reticular formation (Gi, GiV, LPGi; plates 9 to 13) is broader that the parvicellular reticular formation (PCRt; plates 9 to 13). As for other diprotodontids, this would be consistent with a major role for trigeminal somatosensation in analysing the environment and feeding information to the brainstem reticulospinal pathways for co-ordination of forelimb movement. A putative lateral vestibulospinal tract (lvs in plates 11 to 13) is identified and may mediate control of spinal cord motoneurons in response to vestibular input.
Sense organs
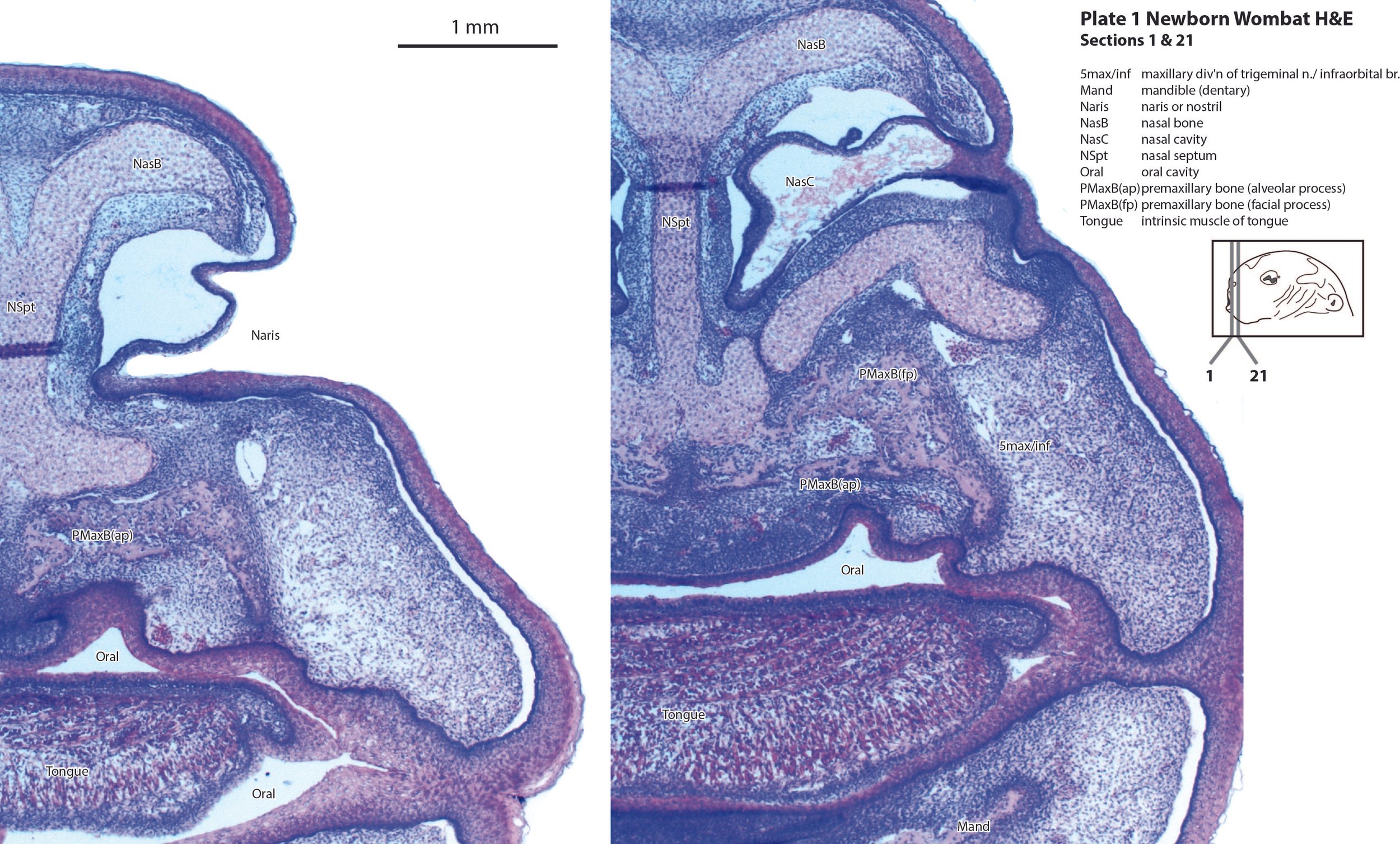
The olfactory epithelium of the nasal cavity (olfepith in plates 2, 3) is thicker than respiratory epithelium (respepith in plates 2, 3) and olfactory nerve fibres (olf in plate 3) emerge from the former. The eye structure is immature and lids closed, with few or no postmitotic neurons, but the sensory retina (Retina in plates 4, 5), and pigment epithelium (Pig in plates 4, 5) can be distinguished. The inner ear has progressed to a stage where cochlear duct (plate 9), utricle (plates 10, 11), saccule (plates 10, 11), semicircular ducts (plates 10 to 12) and endolymphatic duct and sac (plates 11, 12) can be identified. Mystacial vibrissae (Vibr in plate 2) are less mature than in the tammar wallaby and brushtail possum at the same age. This may reflect significant behavioural differences between the newborn wombat and other diprotodontids.
Musculoskeletal system
Cartilaginous models for the premaxilla, nasal, ethmoid, lacrimal, maxillary, presphenoid, basal sphenoid, basal occipital, squamosal, periotic, ex- and supraoccipital bones are present.
Acknowledgements
I would like to thank Dr Peter Giere of the MfN, Berlin Germany, for access to the collection and for all his kind help during the work.