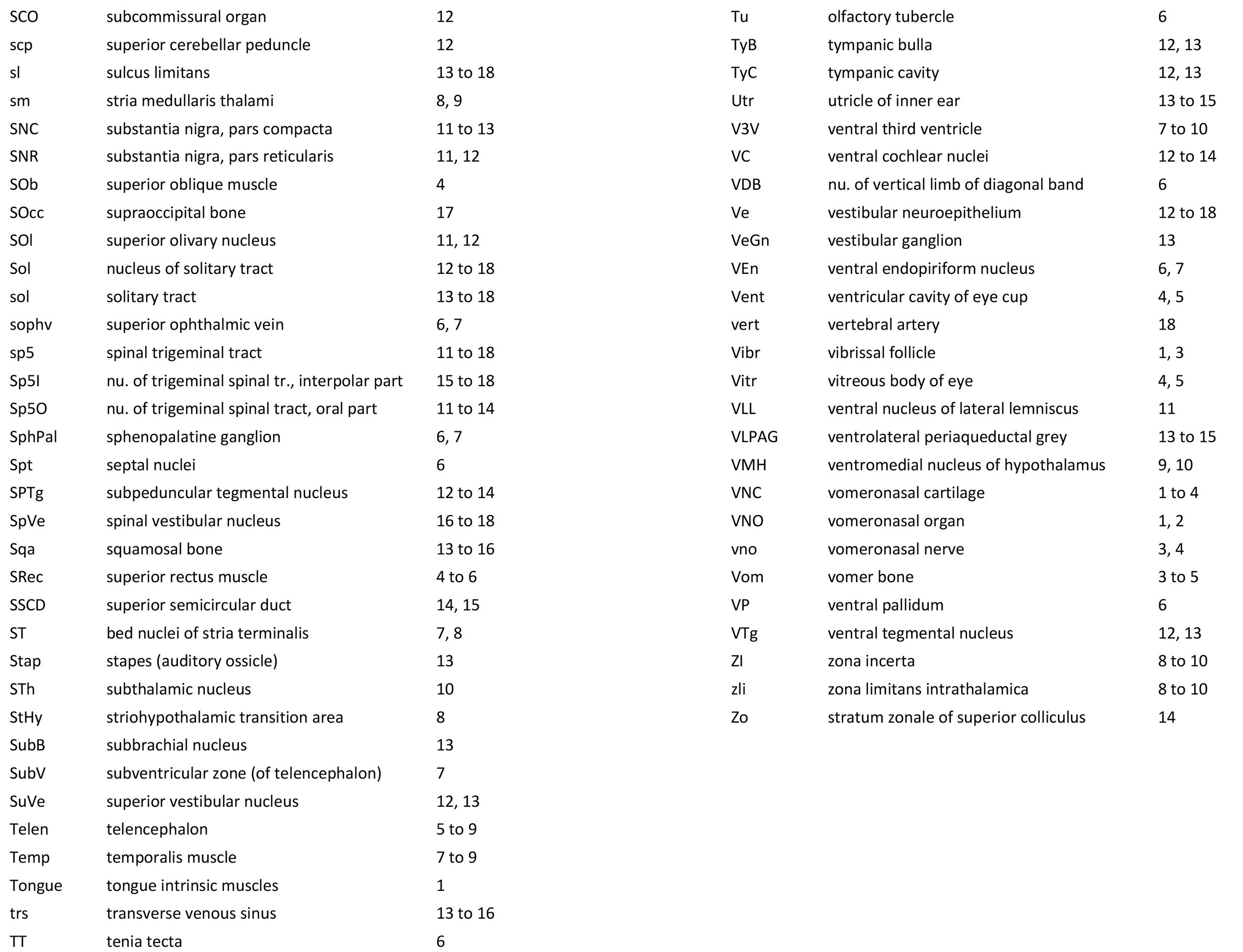
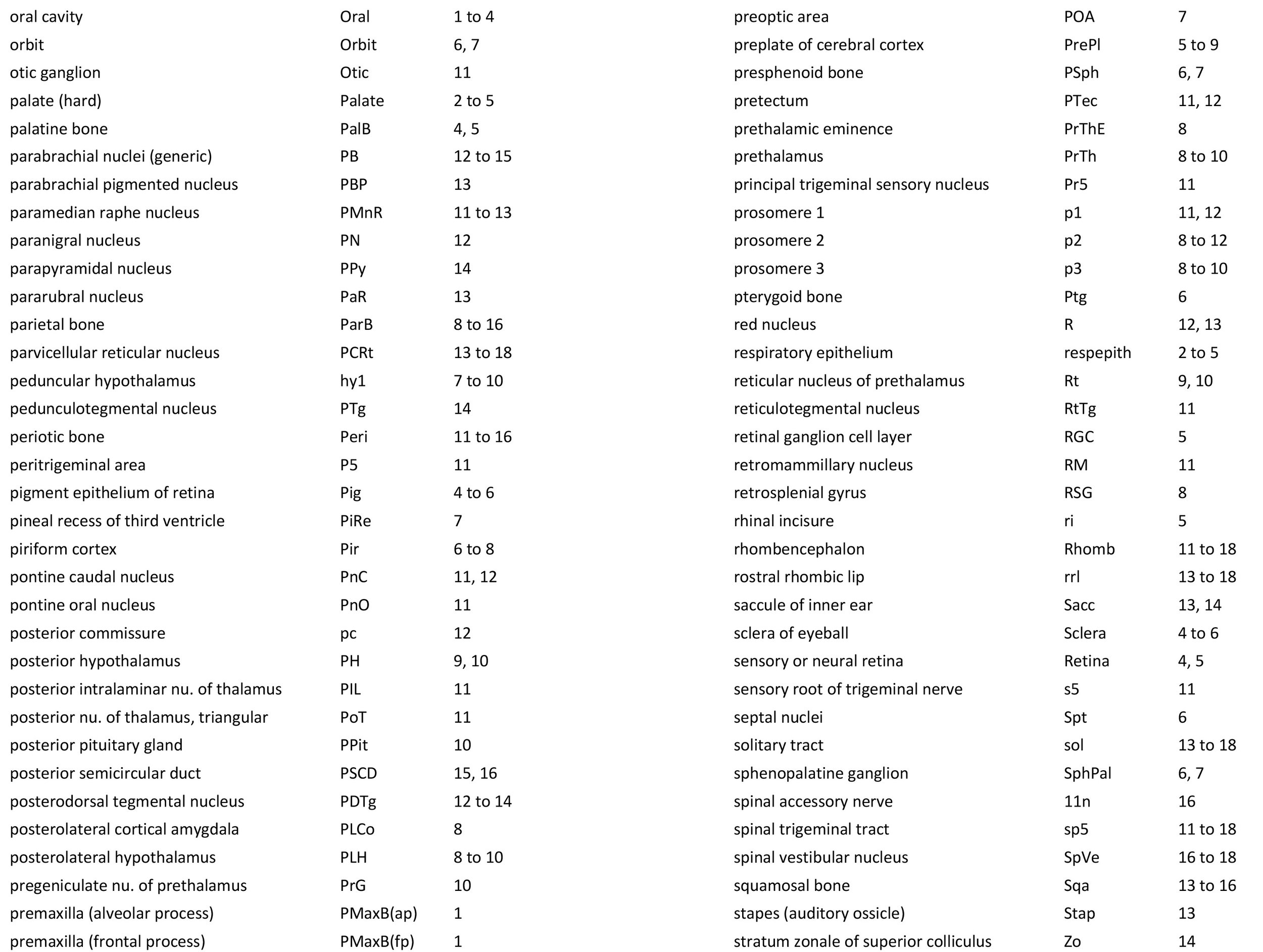
Atlas of the Head and Neck of a Newborn Koala (MS353)
Introduction
The koala (Phascolarctos cinereus) is an arboreal vombatiform diprotodontid marsupial found down the eastern and southern coastal areas of mainland Australia from Queensland to South Australia. Adult koalas can weigh up to 15 kg and reach body lengths of 85 cm. Their usual diet is eucalyptus leaves, which are nutritionally poor and contain toxins (terpenes and phenols). Koalas are highly sedentary, probably due to the low caloric content of their diet. The brain is relatively small for the body mass and the cerebral cortex is smooth (lissencephalic) occupying only 61% of the endocranial volume (Haight and Nelson, 1987; Tyndale Biscoe 2005). This probably allows the koala to conserve energy in response to the poor nutritional quality of its eucalyptus leaf diet (Flannery, 1994; Tyndale-Biscoe, 2005).
Gestation is 33 to 35 days. Koalas give birth from October to May (spring to autumn) and the (usually single) newborn joey weighs approximately 0.5 g, with a greatest length (GL) of 15 to 16 mm. As for other diprotodontid marsupials, the immature newborn must climb to the pouch to continue development. Lactation may continue for as much as a year. The joey reaches 0.5 kg by about 7 months after birth.
Methods
The specimen illustrated here is MS353 (Phascolarctos cinereus, newborn, GL of 16.5 mm) of the Hill collection stored at the Museum für Naturkunde in Berlin. Head length of the specimen after dehydration is approximately 6 mm.
The specimen had been collected in the late 19th century, embedded in paraffin wax and sectioned transversely at 15 µm thickness, before being stained with hematoxylin and eosin.
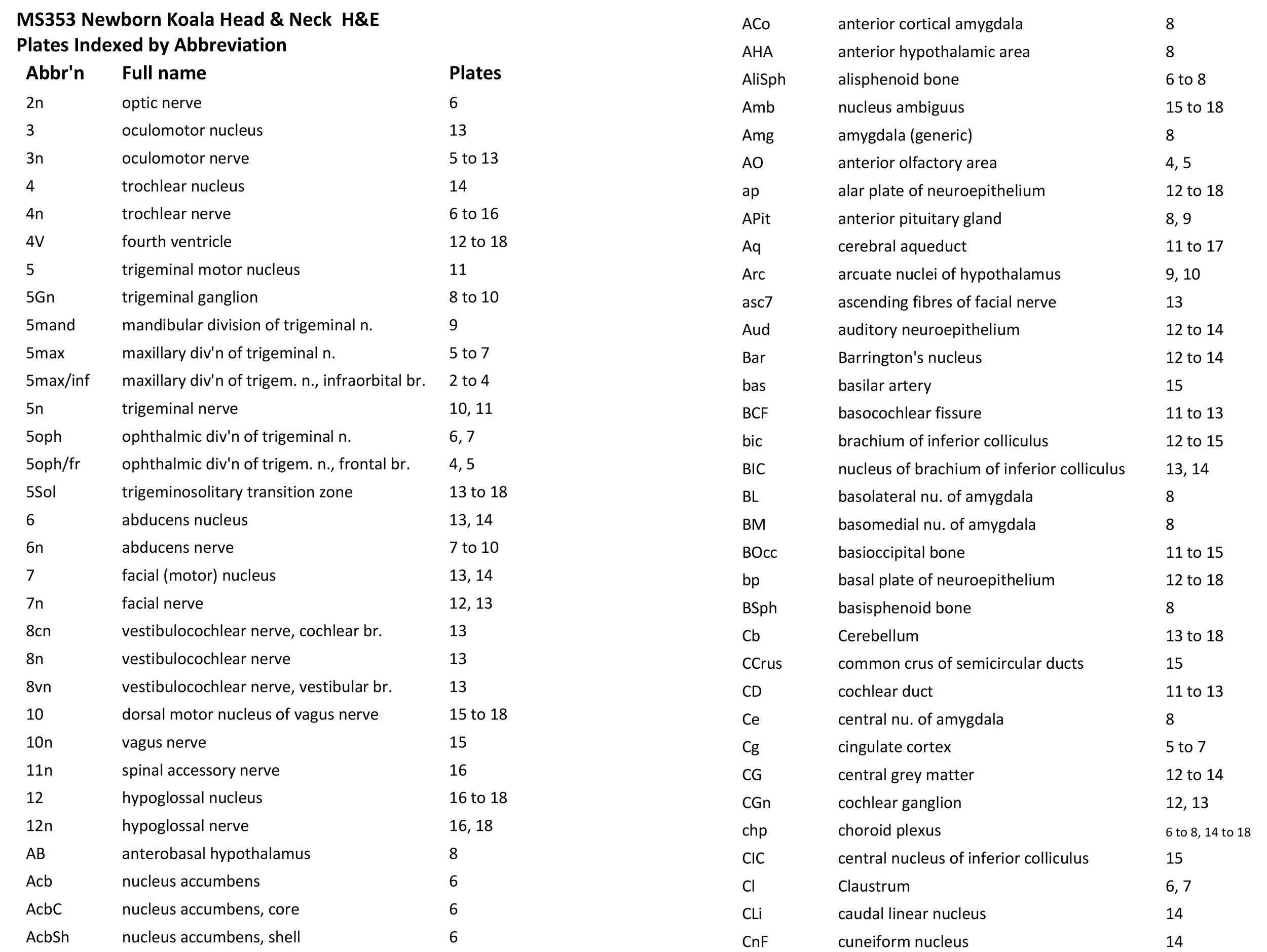
Sections at approximately 150 µm intervals were photographed with the aid of a Zeiss Axioplan2 fitted with an AxioCam MRc5 camera. Sections at 300 µm intervals have been depicted in the plates and the section numbers are consecutive from the snout tip. All images were calibrated by photographing a scale bar at the same magnification. Images were placed in Adobe Illustrator 2021 and delineated. Developmental regions (i.e. neuroepithelium) destined to give rise to adult structures have been denoted by the adult structure’s name with an asterisk (e.g. Cx* denotes the developmental field of the cerebral isocortex).
An image of the profile of a newborn koala head was not available, so a line diagram of the head of a newborn tammar wallaby has been used for the finder illustration indicating section position.
Notes on the specimen
General observations
The oral shield (Schneider and Gurovich, 2017), which is an epidermal thickening anterior to the nasal, premaxilla, maxilla and dentary, is less prominent in this species than in the newborn dasyurid but is represented by some epidermal thickening in section 1, plate 1. In general, the central nervous system, cranial skeleton and craniofacial musculature in the newborn of this species are similar to a newborn macropod (e.g. tammar wallaby).
Central nervous system
As for other diprotodontid marsupials, the central nervous system is immature in this specimen compared to even altricial newborn placental mammals such as the laboratory rat. In fact, the state of forebrain development is most like that in an E15 laboratory rat, about 1 week before birth.
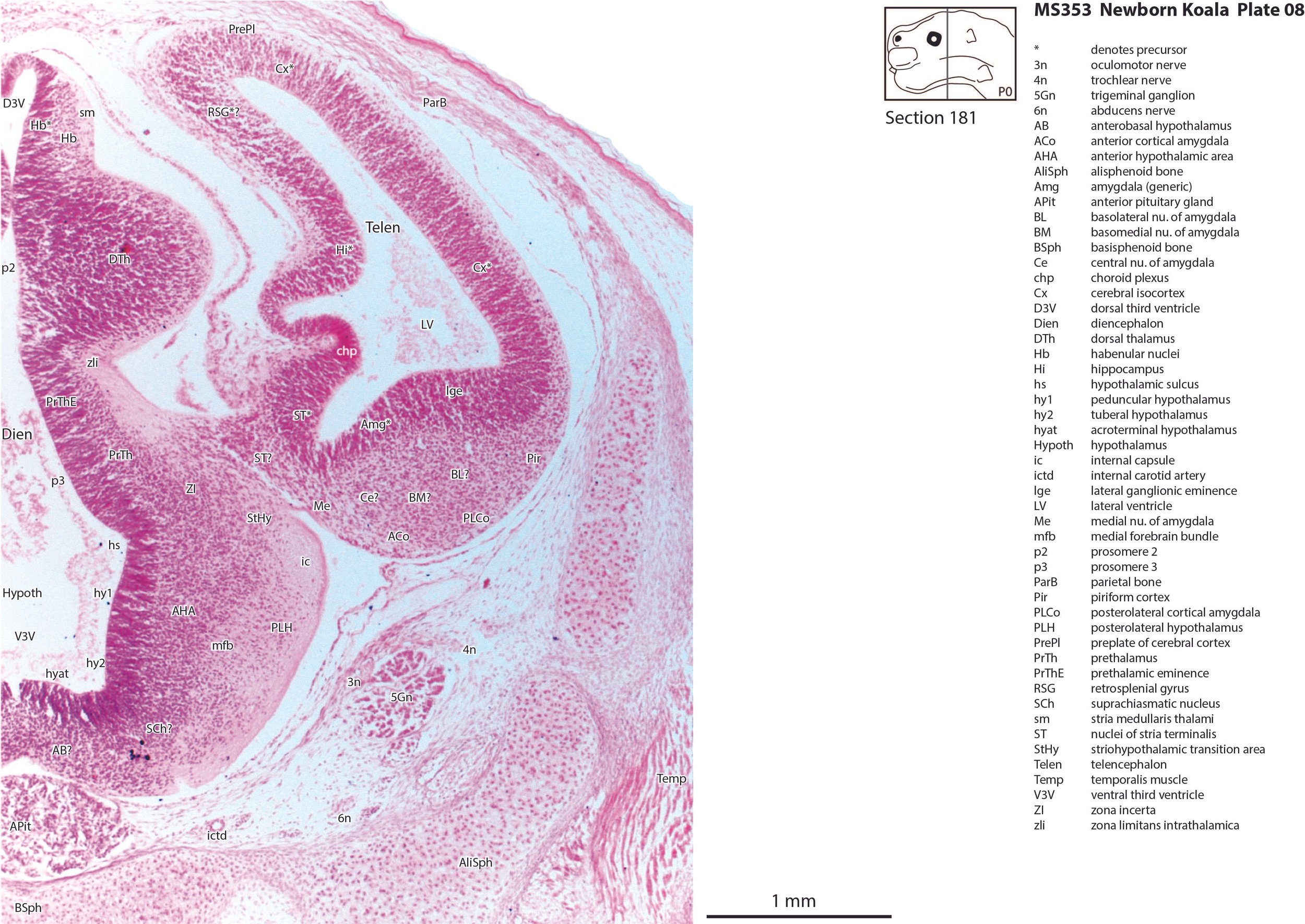
The telencephalon has paired telencephalic vesicles (e.g. plates 5 to 9) but the proliferative compartment known as the ventricular germinal zone (Cx*) is thicker than the preplate (PrePl) that contains the post-mitotic neurons. This is particularly the case in medial parts of the telencephalic vesicle (e.g. plates 6 to 9) where the preplate is negligible. On the other hand, the cortical plate (CxP) is present laterally (see plates 5 to 7). This suggests a distinct neurogenetic gradient in the developing koala isocortex, such that lateral parts (piriform, insula, auditory and lateral somatosensory cortex) are in advance of medial parts (e.g. medial motor and visual cortex).
Some olfactory nerve fibres are visible (olf in plates 3, 4), but very few neurons are present in the olfactory bulbs, with only poor lamination of the developing olfactory bulb (plates 4, 5). Infolding of the hippocampal primordium has begun (plates 6 to 8) but very few neurons have been generated at this stage and no dentate gyrus primordium is visible.
The diencephalon (plates 7 to 12) can be divided into prethalamus (previously ventral thalamus), (dorsal) thalamus, and pretectum, derived from prosomeres 3, 2 and 1, respectively. Some white matter has begun to appear on the exterior of the forebrain. These include the stria medullaris thalami (sm; plates 8 and 9), lateral telencephalic tract (lt; plate 7) and cerebral peduncle (cp; plates 10 to 13). Both the lt and cp are similar in size to those in a newborn tammar, suggesting that a large lt is a common feature of diprotodontid marsupials. The function of the fibres in the lt is unknown, but they may play a role in striatal control of behaviour.
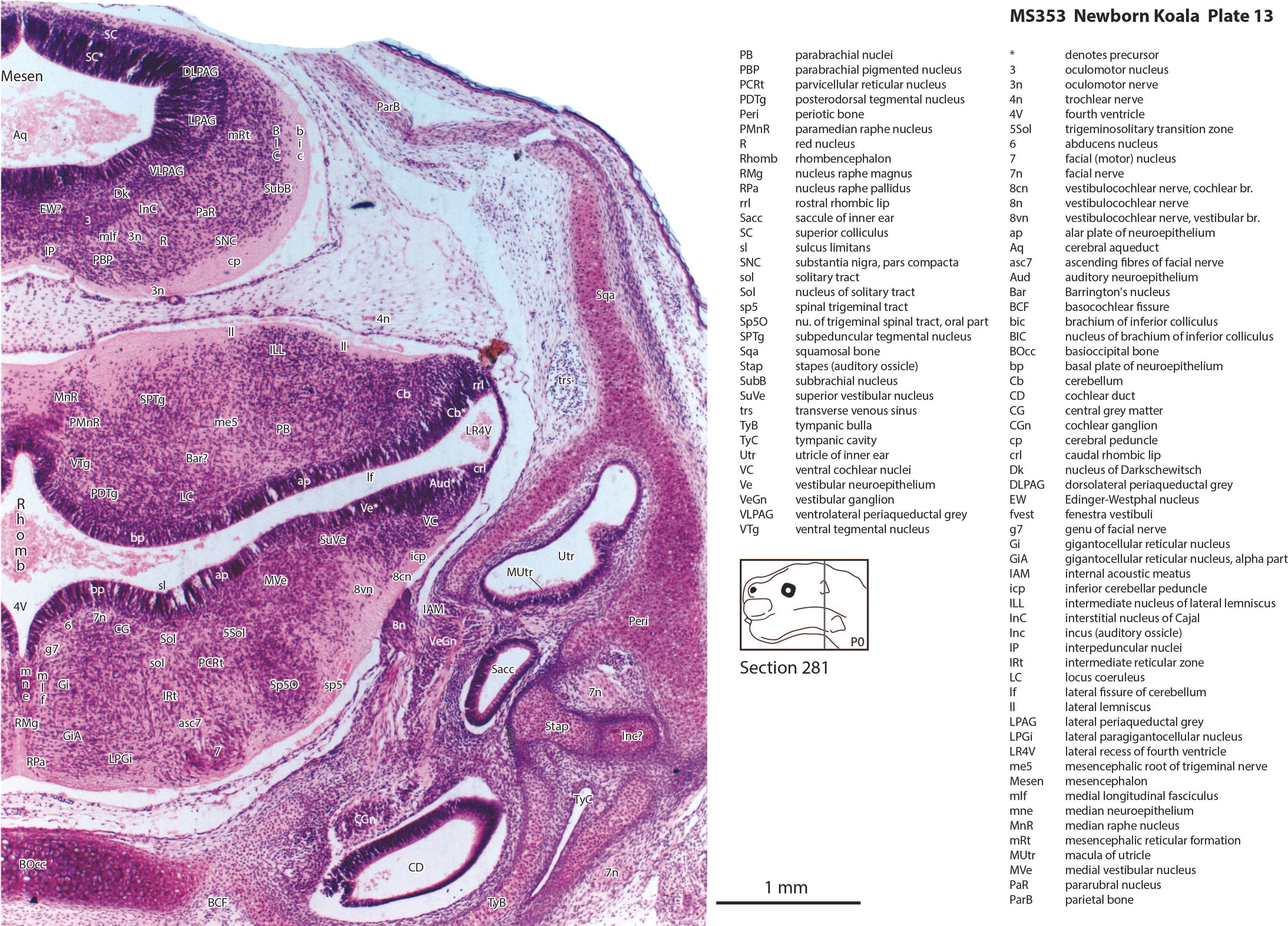
The brainstem of this newborn koala is at a similar level of differentiation to the newborn tammar wallaby. The trigeminal sensory nuclear complex (Pr5, Sp5O, Sp5I, Sp5C; plates 11 to 18) is easily distinguished and the medial magnocellular reticular formation (Gi, GiV, LPGi; plates 12 to 18) is broader that the parvicellular reticular formation (PCRt; plates 13 to 18). This would be consistent with a major role for trigeminal somatosensation in analysing the environment and feeding information to the brainstem reticulospinal pathways for co-ordination of forelimb movement.
The cerebellum is of a similar level of differentiation to an E15/16 rat (plates 13 to 18), with very few postmitotic cells in a nuclear transitory zone (ntz – plate 17) and only rudimentary inferior and superior cerebellar peduncles (plates 12 to 18, and 12, respectively). Precerebellar nuclei are also rudimentary, with only an incomplete inferior olivary nuclear complex (IO in plates 16 to 18) and no pontine nuclei present.
Peripheral nervous system
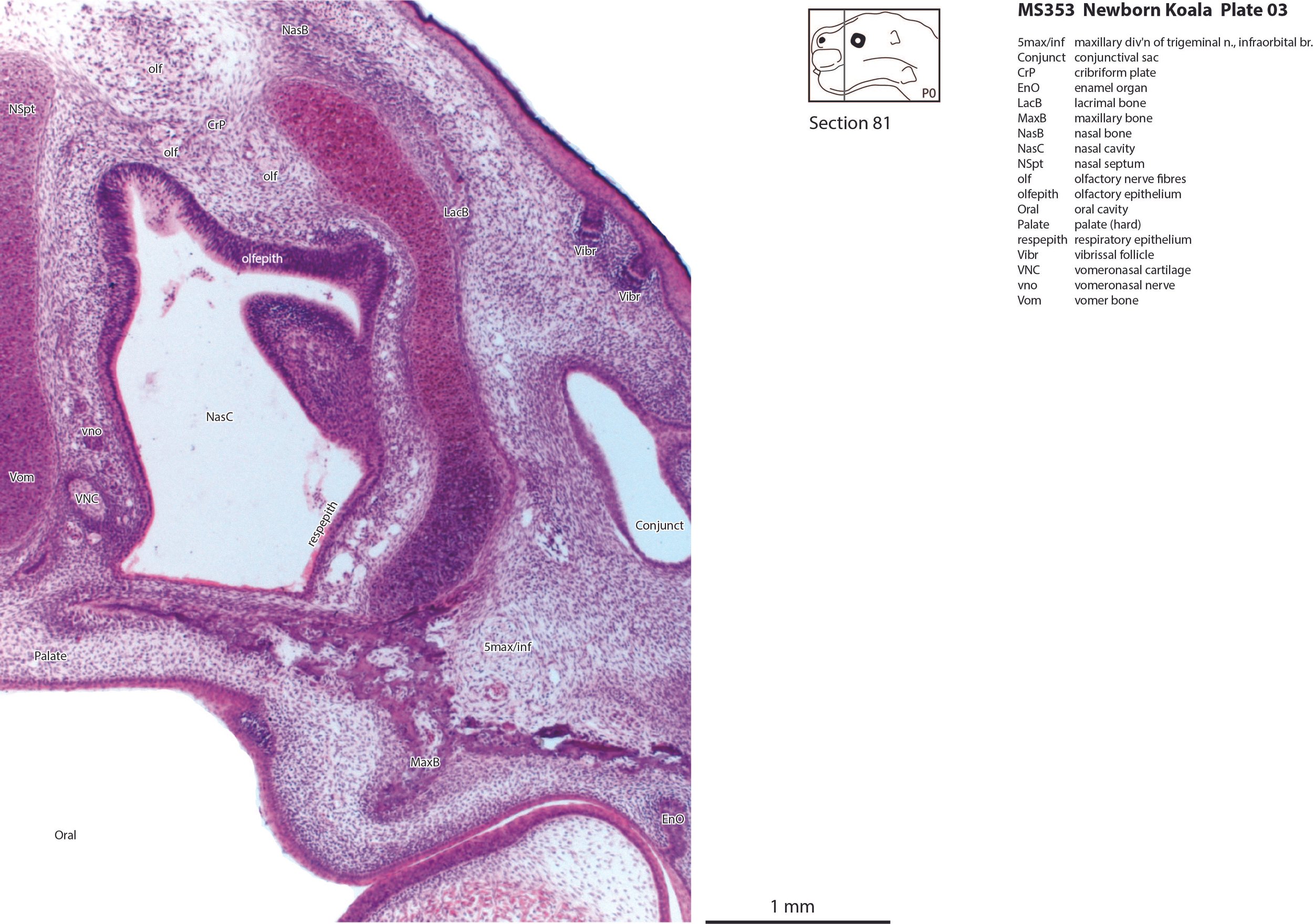
Most cranial nerves are visible (olf in plates 3 and 4, 3n – oculomotor in plates 9 to 15, 4n – trochlear in plates 6 to 16, 5n – trigeminal in plates 10 and 11, 7n – facial in plates 12 and 13, 8n – vestibulocochlear in plate 13, 10n – vagus in plate 15, 11n – spinal accessory in plate 16, 12n – hypoglossal in plates 16 to 18). As with all marsupials, the most differentiated cranial nerve is the trigeminal, with all three divisions visible (ophthalmic, maxillary and mandibular) including some of the major branches of the maxillary and mandibular division (e.g. 5max/inf in plates 2 to 4).
Sense organs
In the nasal cavity, the thicker olfactory epithelium can be distinguished from the thinner respiratory epithelium (plates 1 to 4), but the rest of the olfactory system is still embryonic (see main olfactory bulb discussed above). The lateral olfactory tract (lo in plates 5 to 7) is a good landmark for piriform (primary olfactory) cortex.
The eyes are only rudimentary (plates 4, 5). A conjunctival sac, cornea and lens can be identified, but the optic cup is small. A proliferative compartment in the sensory retina has appeared (plates 4, 5), but very few neurons have been generated. An immature optic nerve (2n) is visible (plate 6).
The Eustachian (pharyngotympanic) tube has just reached the region of the future middle ear (plate 12). The otic vesicle has an endolymphatic duct and sac (plates 15 to 17) visible as an elongated dorsally directed diverticulum. Other subregions (utricle, saccule, cochlear duct and semicircular ducts) are also distinguishable (plates 11 to 15), with sensory maculae visible (MUtr in plates 13, 15; MSacc in plate 14). As regards middle ear bones, a putative stapes is visible (plate 13), along with a cartilaginous model of the incus (plate 13).
Early cutaneous sense organs in the form of mystacial vibrissae (Vibr – plates 1, 3) are seen, but the lingual epithelium does not show any taste buds.
Musculoskeletal system
Cartilaginous models for the premaxilla, nasal, ethmoid, lacrimal, maxillary, basal sphenoid, basal occipital, squamosal, periotic, ex- and supraoccipital bones are present.
Acknowledgements
I would like to thank Dr Peter Giere of the MfN, Berlin Germany, for access to the collection and for all his kind help during the work.
References
Flannery TF (1994) The Future Eaters. Sydney: Reed Books.
Haight JR, Nelson JE (1987) A brain that doesn’t fit its skull: a comparative study of the brain and endocranium of the koala Phascolarctos cinereus (Marsupialia: Phascolarctidae). In Possums and Opossums: Studies in Evolution (ed. M. Archer). Sydney: Surrey Beatty and Sons. pp. 331–352.
Schneider NY, Gurovich Y (2017) Morphology and evolution of the oral shield in marsupial neonates including the newborn monito del monte (Dromiciops gliroides, Marsupialia Microbiotheria) pouch young. J Anat 231, 59-83.
Tyndale-Biscoe H (2005) Life of Marsupials. Collingwood: CSIRO publishing.