Atlas of the Head and Thorax of a Newborn Eastern Quoll (MS133b)
Introduction
The endangered eastern quoll (Dasyurus viverrinus) is a solitary, nocturnal, medium-sized marsupial carnivore (adult males averaging 1.1 kg, adult females averaging 0.7 kg) from Tasmania and occasional sanctuaries on mainland Australia. The gestation period is 19 to 24 days, after which time up to 20 young are born. The young develop in a shallow fur-lined pouch on the maternal abdomen, where there are only 6 to 8 teats, so unattached young die. Pouch development is prolonged, with young remaining attached to the teat for 2 months and weaning at 5 to 6 months.
Methods
The specimen illustrated here is MS133b (Stage A, newborn) of the Hill collection stored at the Museum für Naturkunde in Berlin. The specimen had been collected in the late 19th century, embedded in paraffin wax and sectioned at 10 µm thickness, before being stained with hematoxylin and eosin.
The sections at approximately 100 µm intervals were photographed with the aid of a Zeiss Axioplan2 fitted with an AxioCam MRc5 camera. All images were calibrated by photographing a scale bar at the same magnification. Images were placed in Adobe Illustrator 2021 and delineated. Developmental regions (i.e. neuroepithelium) destined to give rise to adult structures have been denoted by the adult structure’s name with an asterisk (e.g. Cx* denotes the developmental field of the cerebral isocortex).
Notes on the specimen
General observations
The most striking feature of the head is the large oral shield (Schneider and Gurovich, 2017; plates 3 to 6), which is an epidermal thickening anterior to the nasal, premaxilla, maxilla and dentary. The forelimbs are thick and the antebrachium and manus pronated so that the digits can grasp maternal fur as they flex. The anterior neck is covered by a large cervical swelling composed of loose connective tissue.
Central nervous system
The central nervous system is very immature in this specimen. The telencephalon has only rudimentary telencephalic vesicles (plate 1c) and only a single ganglionic eminence can be seen (see plate 2). Some olfactory nerve fibres are visible, but very few neurons are present in the olfactory bulbs, with only an olfactory nerve fibre layer and a very primitive undifferentiated cellular layer visible (plate 1b, c).
The diencephalon (plates 2, 3) can be divided into prethalamus, (dorsal) thalamus, and pretectum. There is very little white matter at any levels of the forebrain (plates 2 to 4) indicating that very few forebrain pathways have been laid down at this stage.
The largest groups of neurons are present in the rostral and midlevel brainstem in the lateral sensory column (principal trigeminal sensory nucleus and nucleus of trigeminal spinal tract; plates 4 to 6) and the medial gigantocellular reticular formation (Gi). This would be consistent with a major role for trigeminal somatosensation in analysing the environment and feeding information to the brainstem reticulospinal pathways for co-ordination of forelimb movement.
The cerebellum is only rudimentary (plate 4), with very few postmitotic cells and no apparent connections with the brainstem.
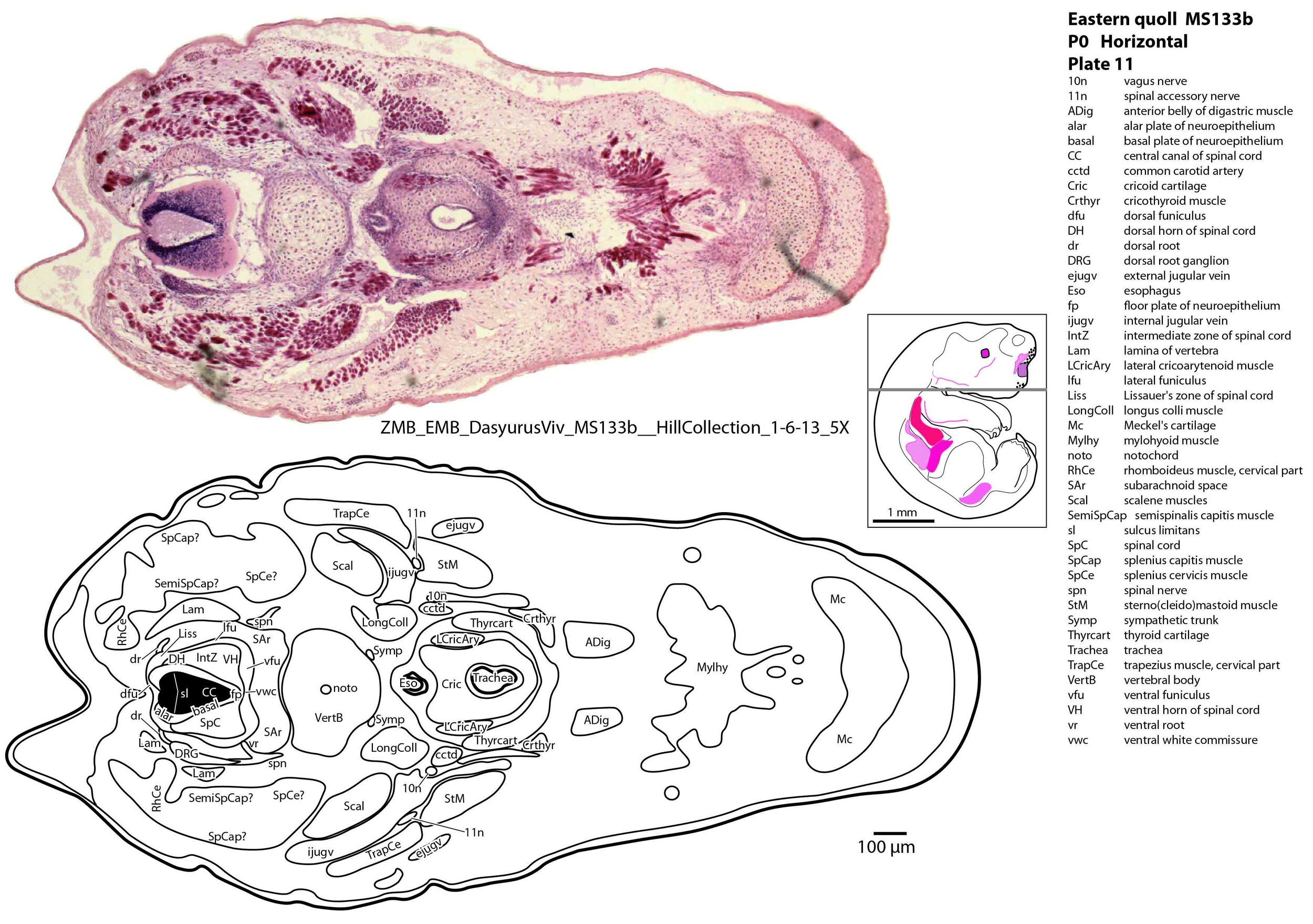
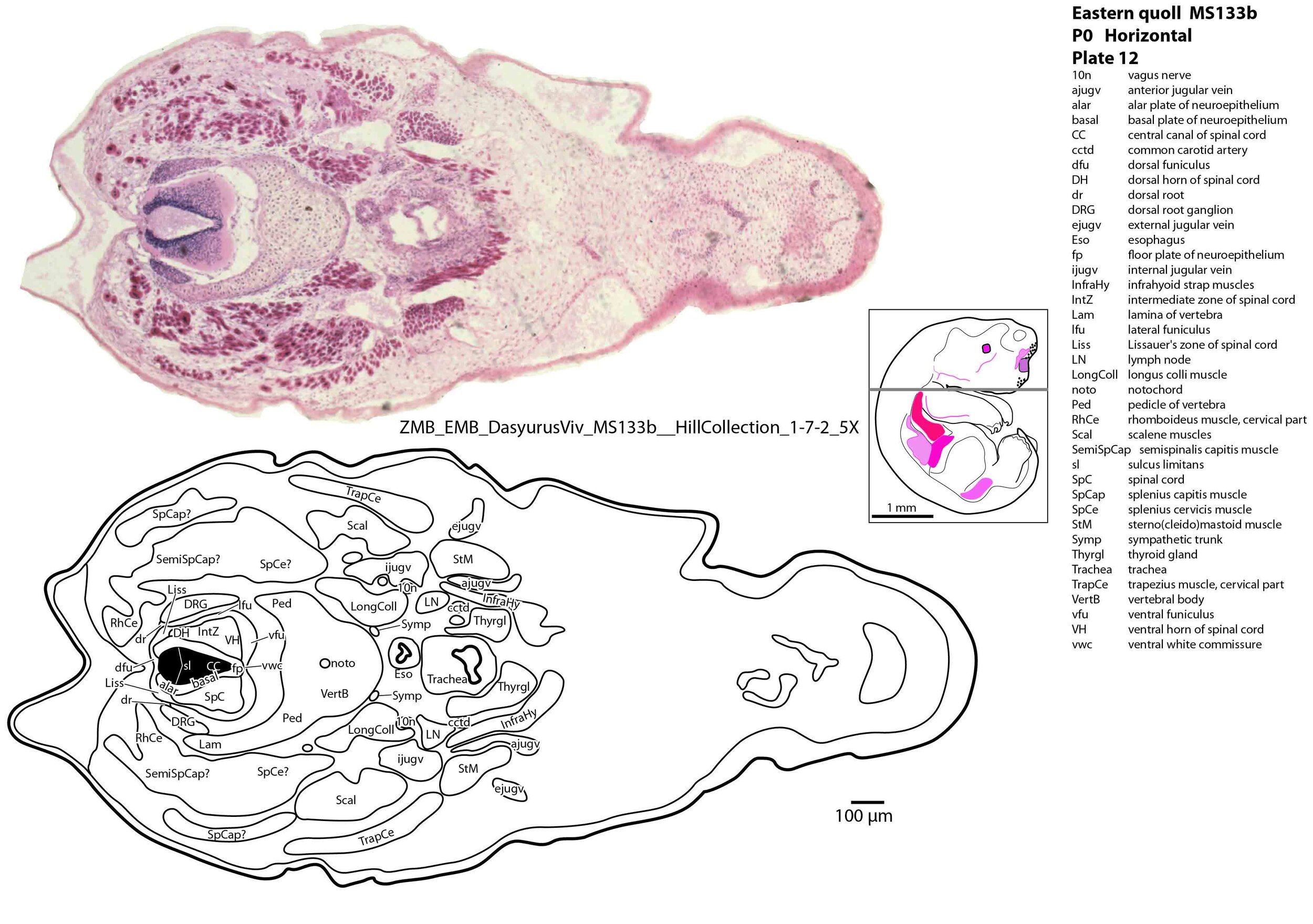
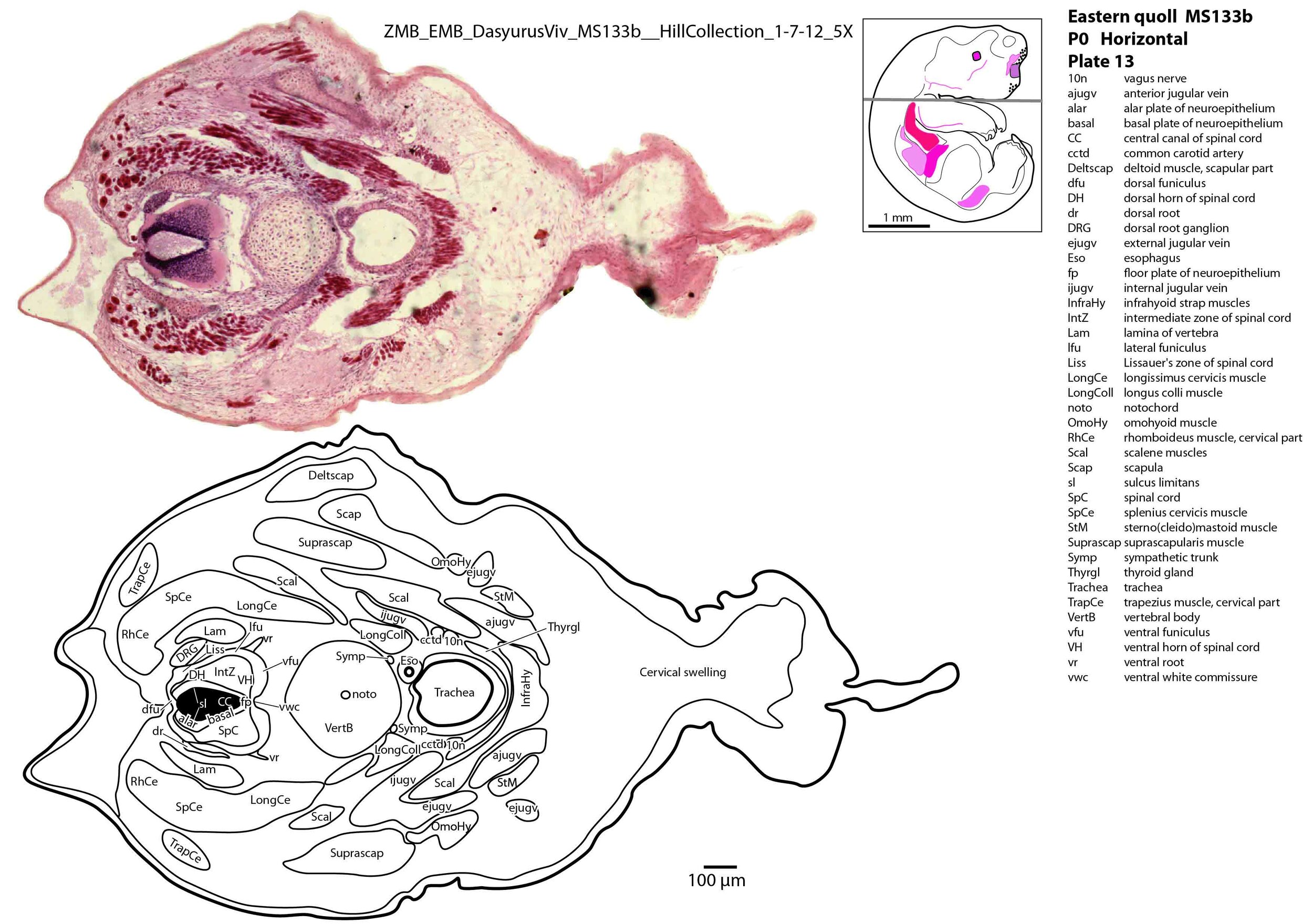
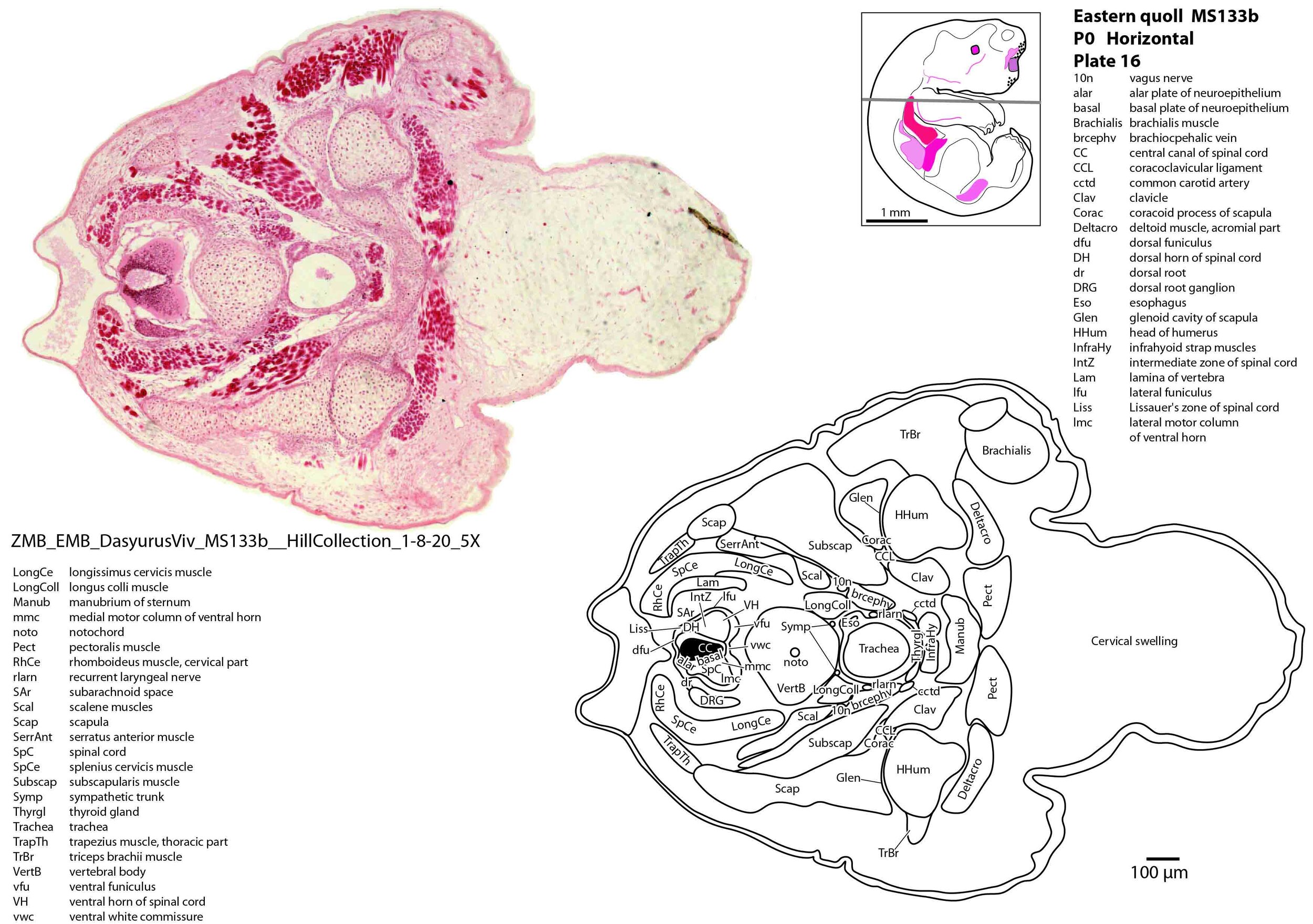
The spinal cord is most differentiated in the cervical region (e.g. plate 14), but even here there is very limited differentiation of the dorsal (sensory) horn. By contrast the ventral (motor) horn is large and broad in the lower cervical region with emergence of medial and lateral motor columns for control of axial and appendicular muscles, respectively. The morphogenetically important floor plate is a consistent feature throughout the spinal cord, but the roof plate is thin and discontinuous, particularly in the upper and mid cervical levels (plate 12) where rostral neuropore closure may have recently occurred.
Peripheral nervous system
Only a few cranial nerves are clearly visible (CN1 – olfactory, CN5 – trigeminal, CN7 – facial, CN10 – vagal, CN11 – spinal accessory, CN12 - hypoglossal). No eye muscle nerves could be seen. The most differentiated cranial nerve is the trigeminal, with all three divisions visible (ophthalmic, maxillary and mandibular) including some of the major branches of the mandibular division (e.g. ling and ialvn in plate 8).
The brachial plexus nerves are prominent crossing over the first rib (plate 18), with at least the radial and median nerves clearly visible in the brachium (plate 22).
Sense organs
In the nasal cavity, the thicker olfactory epithelium can be distinguished from the thinner respiratory epithelium (plate 1c), but the rest of the olfactory system is still embryonic.
The eyes are only rudimentary (plates 3, 4). A conjunctival sac and cornea can be identified, but the lens vesicle and optic cup are tiny and primitive. An optic stalk can be traced to the hypothalamus, but there is no optic nerve visible.
The external acoustic meatus is uncanalised (plates 7, 8) and the Eustachian tube has not reached the region of the future middle ear (plate 8). The otic vesicle has an endolymphatic duct (plate 6) visible as a small dorsally directed diverticulum, but other subregions (utricle, saccule, and cochlear duct) are not clearly separated (plate 6). A putative stapes is visible (plate 7), but no other auditory ossicles, which presumably are still within the arch 1 mesenchyme (plate 7).
No cutaneous sense organs are seen and the lingual epithelium does not show any taste buds.
Musculoskeletal system
Cartilaginous models for the premaxilla, nasal, maxillary, basal sphenoid, basal occipital and squamosal bones are present (plates 1 to 7). The mandible (dentary) is developing around the Meckel’s cartilage (Plates 7 to 11).
Muscles have been identified by reference to Diogo et al. (2016) and Warburton and Marchal (2017). No facial muscles are visible, but rudimentary pterygoid (plate 7) and masseteric (plate 6) masticatory muscles are present. Intrinsic and extrinsic tongue musculature is present (plates 8 to 10), along with stylopharyngeus and salpingopharyngeus muscles inserting into the pharyngeal tube (plate 9). Two intrinsic laryngeal muscles (lateral cricoarytenoid and cricothyroid) are visible (plate 11).
Vertebrae are composed of a large body, with pedicles and laminae visible (e.g. plat 15), but the neural arch is incomplete. Post-, para and prevertebral muscles are present as groups, with some specific muscles (scalenes, trapezius, sternocleidomastoid, longus colli) distinguishable (plates 6 to 27).
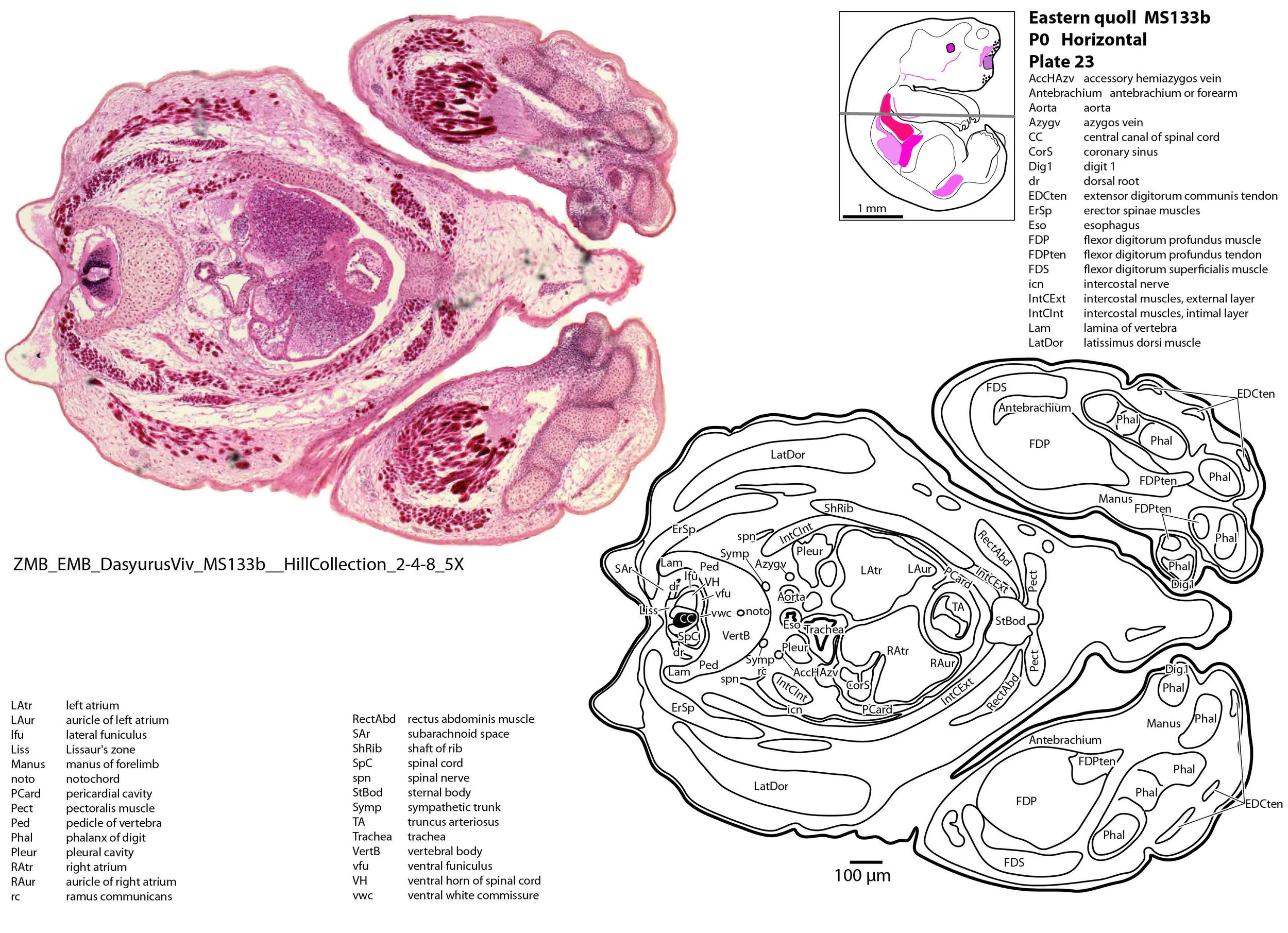
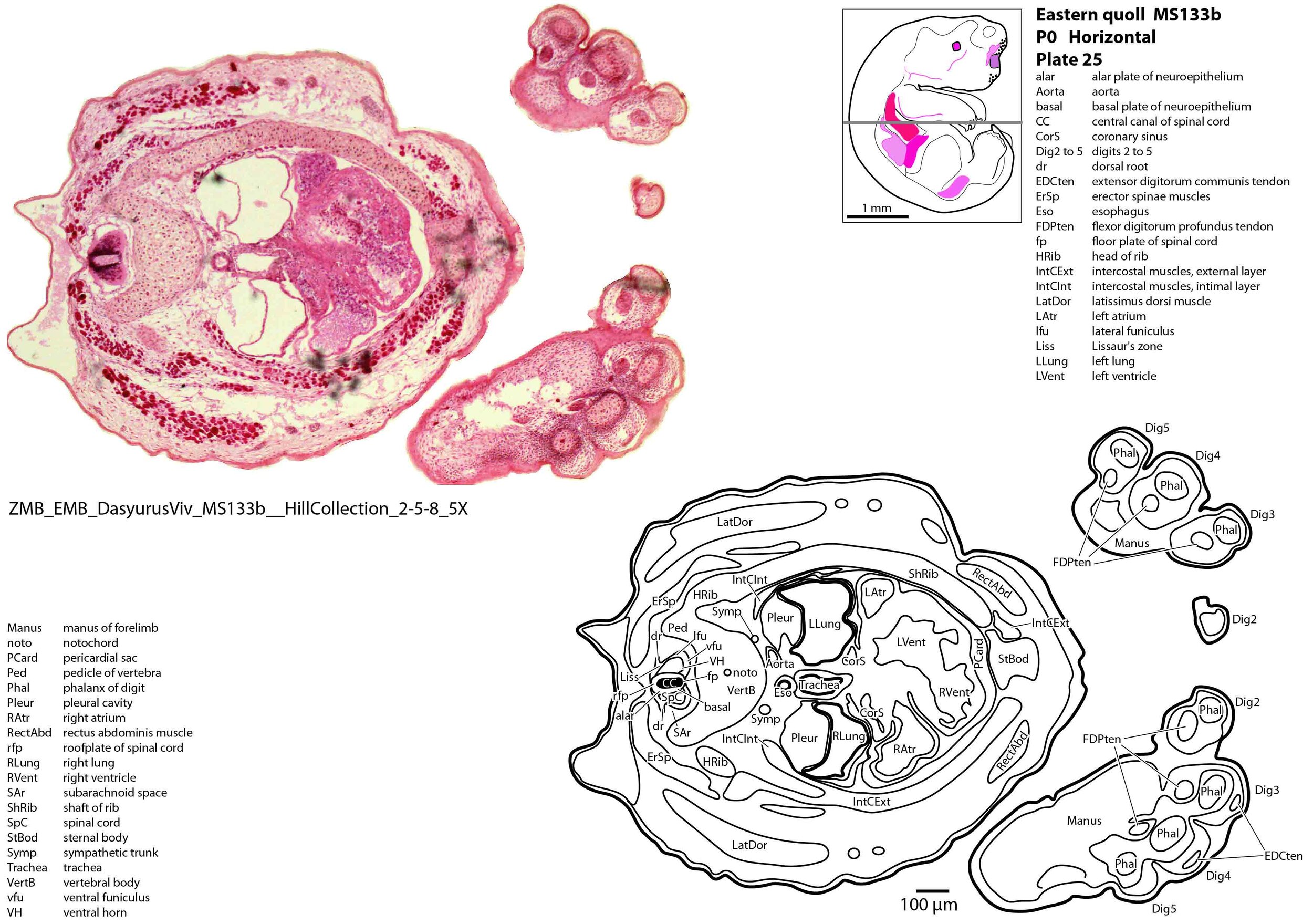
The forelimb skeleton is robust but unmineralised (see thick cartilaginous models of the scapula, clavicle, humerus, radius and ulna, carpals, phalanges). Large forelimb muscle groups required for locomotion (pectoralis, latissimus dorsi, biceps brachii, brachialis, triceps brachii) and grasping/digit extension (flexor digitorum profundus, extensor digitorum communis) are visible (plates 19 to 26)(Warburton and Marchal, 2017).
The chest wall is composed of a manubrium sterni, sternal body, ribs and thoracic vertebrae. Intercostal muscles can be divided into an external layer outside the neurovascular bundle (i.e. the intercostal nerve and accompanying intercostal vessels), and an intimal layer deep to the neurovascular bundle (see plate 24).
Gastrointestinal tract
The gastrointestinal tract of this specimen is empty of milk, suggesting that attachment to the teat and commencement of suckling was not completed at the time of death. Only one set of enamel organs (tooth primordia) is visible in the maxilla (plate 5) and none in the dentary.
The esophagus is very small (external diameter 60 to 80 µm, internal diameter 10 to 40 µm) and attached to the left dorsal side of the much larger trachea at the level of the thoracic inlet (see plate 16).
Respiratory system
The nares are large rostrally and lead to the nasopharynx. The laryngeal inlet (plate 9) is bounded by the epiglottis and aryepiglottic folds and elevated above the level of the soft palate so that ventilation can proceed while the joey is suckling and milk flows through the laryngopharynx. Laryngeal cartilages (epiglottis, thyroid, cricoid, arytenoid) are all present.
The trachea is very large (internal diameter 200 to 250 µm) and occupies much of the thoracic inlet (plate 16). The trachea opens into main bronchi which expand into unseptated lungs (Ferner, 2021; plates 25 to 27).
Cardiovascular system
The atria of the heart are incompletely divided (plate 24), with a communication between left and right sides allowing mixing of blood as an adaptation for skin respiration (Ferner, 2020). The right atrium receives blood from an inferior vena cava and paired brachiocephalic veins. Pulmonary veins draining into the left atrium are very small and difficult to identify.
The ventricles are also incompletely divided (plate 25), with only a partial interventricular septum. The outflow tract from the two ventricles is also undivided at the level of the bulbus cordis (plate 24) and truncus arteriosus (plate 23), meaning that mixing of blood occurs at multiple sites within the heart.
Acknowledgements
I would like to thank Dr Peter Giere of the MfN, Berlin Germany, for access to the collection and for all his kind help during the work.
References
Diogo R, Bello-Hellegouarch G, Kohlsdorf T, Esteve-Altava B, Molnar JL (2016) Comparative myology and evolution of marsupials and other vertebrates with notes on complexity, bauplan, and “scala naturae”. Anat Rec 299, 1224-1255.
Ferner K (2020) Development of the skin in the eastern quoll (Dasyurus viverrinus) with focus on cutaneous gas exchange in the early postnatal period. J Anat 238, 426-445.
Ferner K (2021) early postnatal lung development in the eastern quoll (Dasyurus viverrinus) Anat Rec 304, 2823-2840.
Schneider NY, Gurovich Y (2017) Morphology and evolution of the oral shield in marsupial neonates including the newborn monito del monte (Dromiciops gliroides, Marsupialia Microbiotheria) pouch young. J Anat 231, 59-83.
Warburton NM, Marchal C-R (2017) Forelimb myology of carnivorous marsupials (Marsupialia: Dasyuridae): Implications for the ancestral body plan of the Australidelphia. Anat Rec 300, 1589-1608.