Introduction to Neurobiology of Australian Marsupials
What is Neurobiology and why do we want to study and learn about Marsupial brains? Australasian and South American marsupials have much to offer in broadening our understanding of brain function. Both Charles Darwin and J. I. Johnson recognised that the native Australian mammals represent a ‘second creation’ or, to couch this in more modern terms, a parallel adaptive radiation. Australian marsupials have made a unique journey through the Tertiary, evolving nervous systems to deal with the special features of the changing Australian environment. For much (but not all) of that time, Australian marsupials were completely isolated from eutherian mammals and many of the megafaunal species of the Pleistocene and recent times evolved large and complex brains. This raises many questions.
Have marsupials used the same neural solutions to meet the demands of their environment as their distant eutherian relative , or have they explored novel adaptations? How does the dramatically different developmental timetable of the marsupial lifestyle influence brain development and adult structure? Does the marsupial brain mediate sexual behaviour in the same way as the eutherian brain? These are some of the abiding questions of marsupial neurobiology.
The term Australian is used to include all those parts of Greater Australia to the east and south of the Wallace line (i.e. including New Guinea and its associated islands, main- land Australia and Tasmania). This corresponds to the continental land mass of Greater Australia (Sahul) as it was at the height of the last glaciation of the Pleistocene, when sea- levels were at their lowest and Australian marsupials could move relatively freely between the Australian mainland and surrounding islands.
Marsupial Evolution:
Marsupial Classification:
Australian Marsupials:
Adult Marsupial Brains:
External morphology of marsupial brains (adults):
The external features of different adult marsupials brains demonstrates that there are visual differences in the relative size of the olfactory bulbs and cortical surfaces in the different marsupial families (Ashwell, 2010).
Olfactory structures: As a proportion of total brain size, the olfactory structures are largest in dasyuridae, notoryctidae and peramelidae. However in the Diprotodonts (kangaroos, koalas, wombats etc) the olfactory lobes decrease in relative size or get smaller with increased overall brain volume.
Gyrification of the cortical surface: Gyrification is the folding of the cerebral cortex. The degree of folding present on the cortical surface varies substantially between species. Some species, such as small dasyurids (e.g. Antechinus swainsonii), have quite smooth (lissencephalic) cortical surfaces, whereas other species, like the eastern grey kangaroo (Macropus giganteus), have highly folded (gyrencephalic) cerebral cortex. As in eutherian mammals (all mammals except monotreme and marsupials), gyrification is more extensive in those species with larger brains and higher body weight.
Brain morphology types Australasian marsupials:
Haight and Murray (1981) have argued that, with respect to gross morphology, the brains of Australian marsupials fall into three types.
Brain morphology Type I: The first type (Type I ) is characteristic of Australian polyprotodonts marsupials Dasyuromorphia and include the family Dasyuridae e.g. Eastern quoll and Peramelemorphia and include the families Peramelidae (Bandicoots) and Thlacomyidae ( Bilby). The brains of these marsupials have a trapezoid outline when viewed from above, with large olfactory bulbs, a linear profile when viewed from the side, and little or no dorsoventral flexion. The degree of foliation or folding of the cerebellar cortex may vary, being extensive in dasyurids and less so in bandicoots.
Brain morphology Type II: The second type of brain is found in non-macropod members of Diprotodontia (Vombatidae, Petauridae and Phalangeridae). Viewed from above they exhibit a more ovoid shape than the brains of polyprotodonts. This is due to the lateral expansion of the parietofrontal cortex in type II brains (Ashwell, 2010). Compared to Type I brains the olfactory bulbs are relatively small and the olfactory cortical regions are correspondingly less extensive. These Type II brains are more rounded dorsally because of a slight dorsoventral flexion and the volumetric proportion of isocortex is greater (Ashwell, 2010).
Brain morphology Type III: The third group of brains (Type III) is characteristic of macropods ( Diprotodontidae: Family Macropodidae) e.g. Potorous tridactylus, Thylogale billardierii, Macropus giganteus and Brush Tailed Rock wallaby. When viewed from above the hemispheres are triangular in outline. Potoroos may have a more trapezoidal shape because of blunting of the frontal pole. However all macropod brains exhibit a pronounced flexion due to the elevation of the rostral (anterior) part of the skull.
Brain morphology Type IV: The bizarre brain of the marsupial mole ( Family Notoryctidae) was not studied by Haight and Murray (1981) however Ashwell (2010) observed that this brain morphology would constitute a fourth type, with exaggeration of the olfactory structures , no dorsoventral flexion of the forebrain and a poorly differentiated cerebellum.
Brain morphology types American marsupials:
South American marsupials also have similar brain types to those found in Australasian marsupials.
Brain morphology Type I: Monodelphis and Didelphis have Type 1 brains. The living Dromiciops gliroides (Microbiotheria) shares similar external brain morphology with Type 1 Australian marsupials.
Brain morphology Type IV: Some caenolestid marsupials such as Lestoros inca (raton rancho) and Canenolestes obscures have similar brain morphology to the marsupial mole (Type IV) (based on illustrations in Ziehen 1897; Herrick 1921; Obenchain 1925).
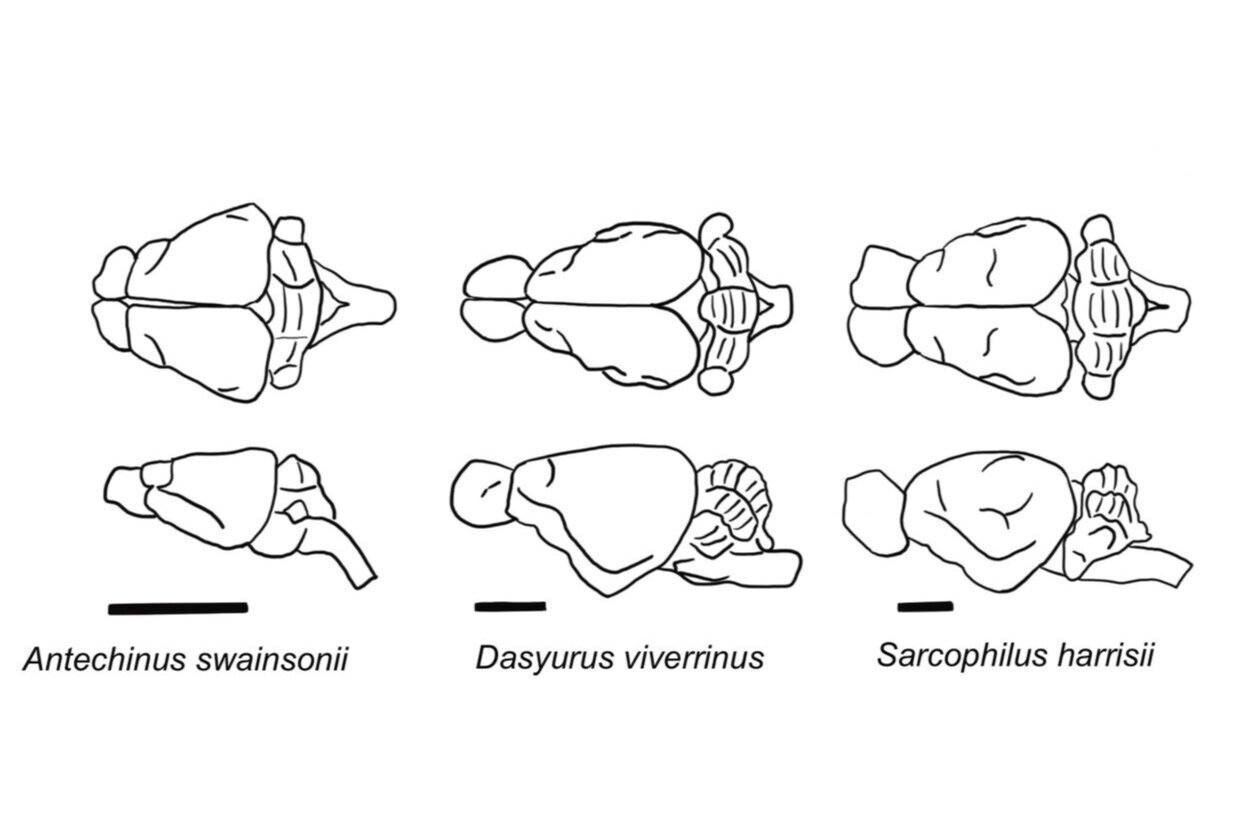
Type I: Dasyurid brains follow type 1 morphology with a trapezoid outline and large olfactory bulbs. Scale bars indicate 10 mm (Modified from Ashwell, 2010).

Type I: Peramelemorphia brains
Type 1 marsupial brain morphology when viewed from above (dorsal or superior view) have a trapezoid outline and large olfactory bulbs. Examples of these brains are bandicoots (Peramelemorphia). Scale bars indicate 10 mm (Modified from Ashwell, 2010).

Type II Diprotodontidae (Non-macropod) brains
The second type of brain is found in non-macropod members of Diprotodontia (Vombatidae- wombats, Petauridae-possums and Phalangeridae). Viewed from above they exhibit a more ovoid shape than the brains of polyprotodonts. This is due to the lateral expansion of the parietofrontal cortex in type II brains (Ashwell. 2010). Scale bars indicate 10 mm (Modified from Ashwell, 2010).
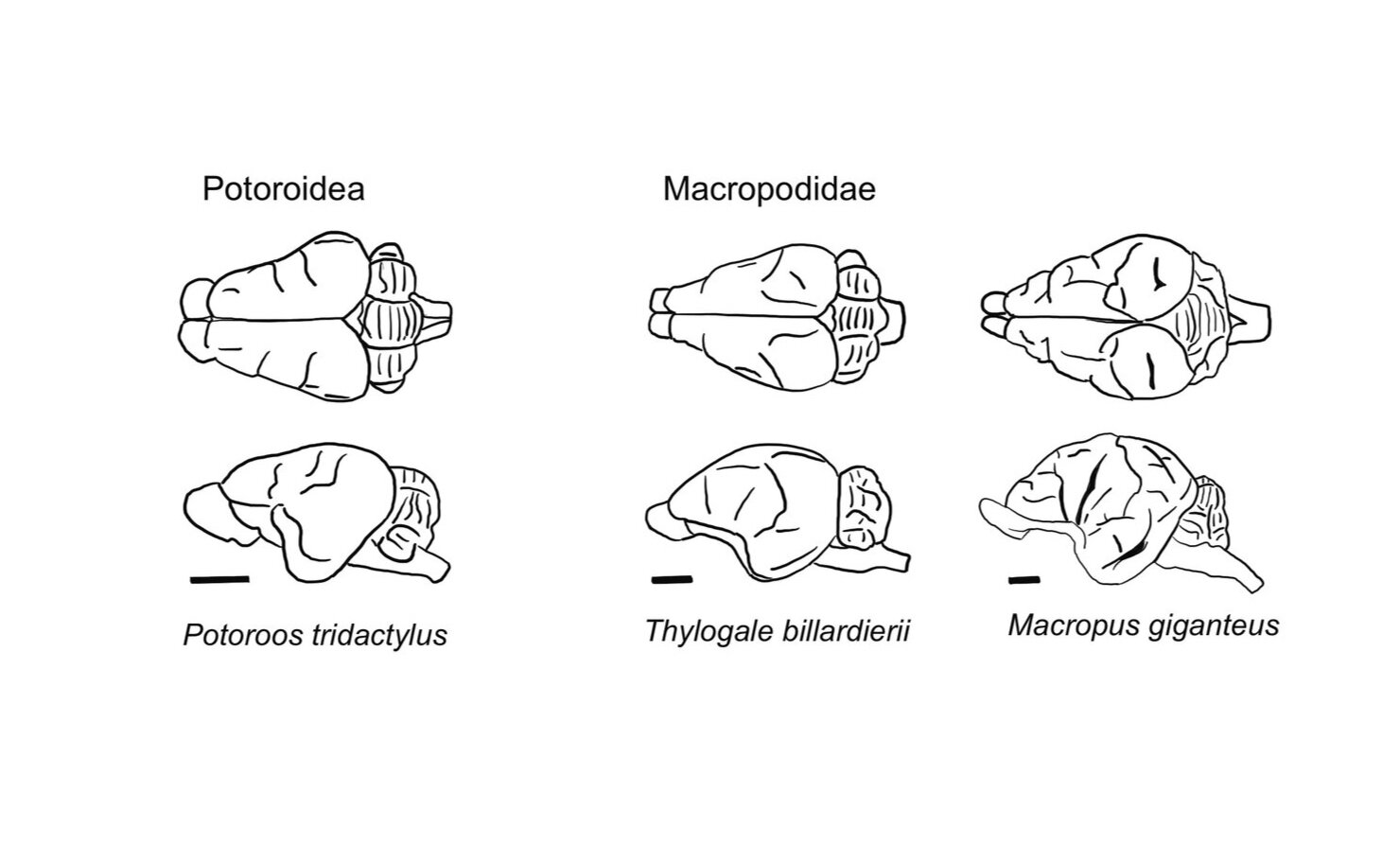
Type III Diprotodontidae (Macropod) brains
The third group of brains (Type III) is characteristic of macropods (Diprotodontidae: Family Macropodidae) e.g. Potorous tridactylus, Thylogale billardierii, Macropus giganteus and Brush Tailed Rock wallaby.

Brain morphology Type IV: The bizarre brain of the marsupial mole ( Family Notoryctidae) was not studied by Haight and Murray (1981) however Ashwell (2010) observed that this brain morphology would constitute a fourth type, with exaggeration of the olfactory structures , no dorsoventral flexion of the forebrain and a poorly differentiated cerebellum.
Inferior view of marsupial brains
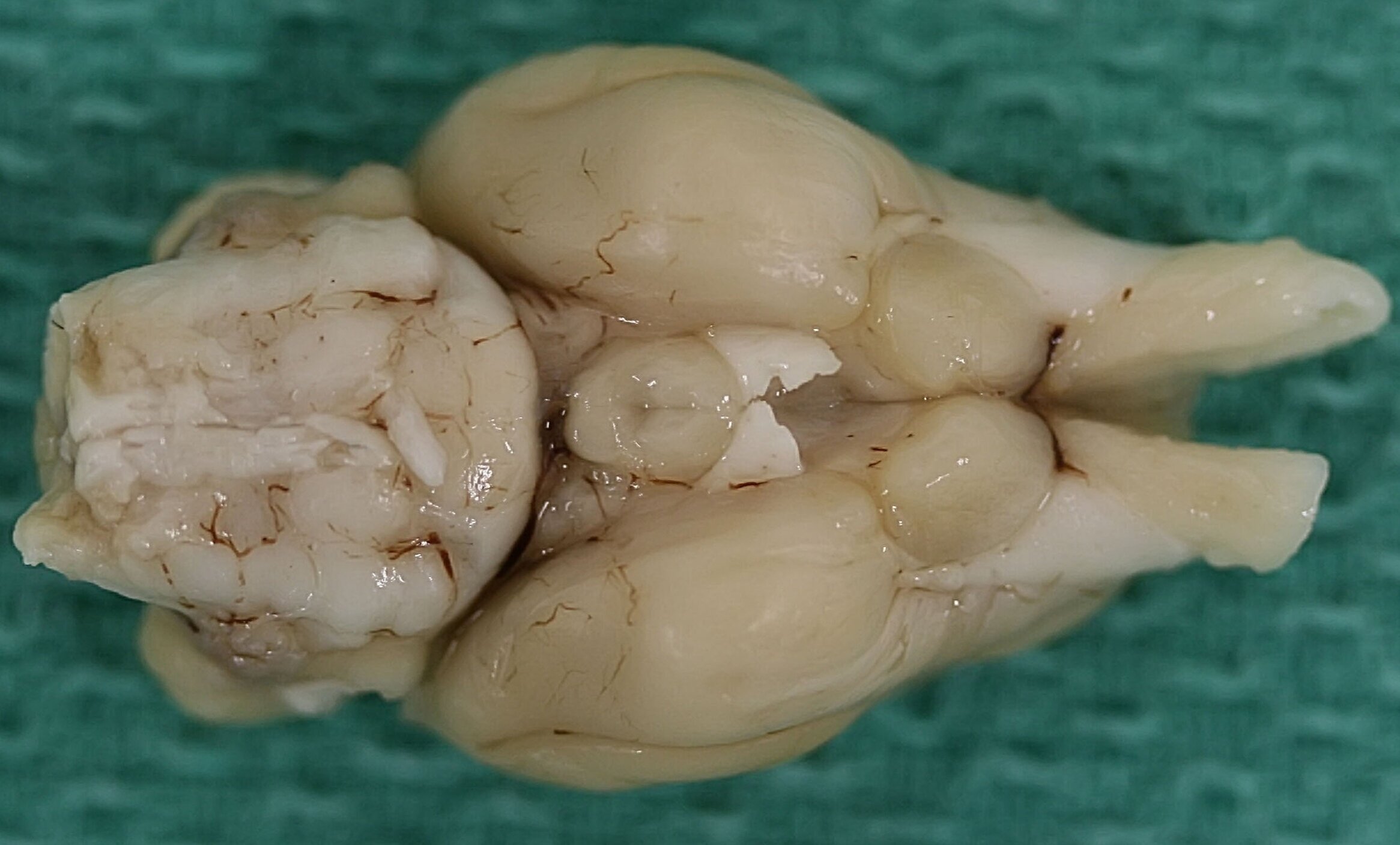
Eastern Quoll Brain inferior view

Brush tail wallaby inferior view

Tasmanian Tiger inferior view

Yellow belly Glider inferior view