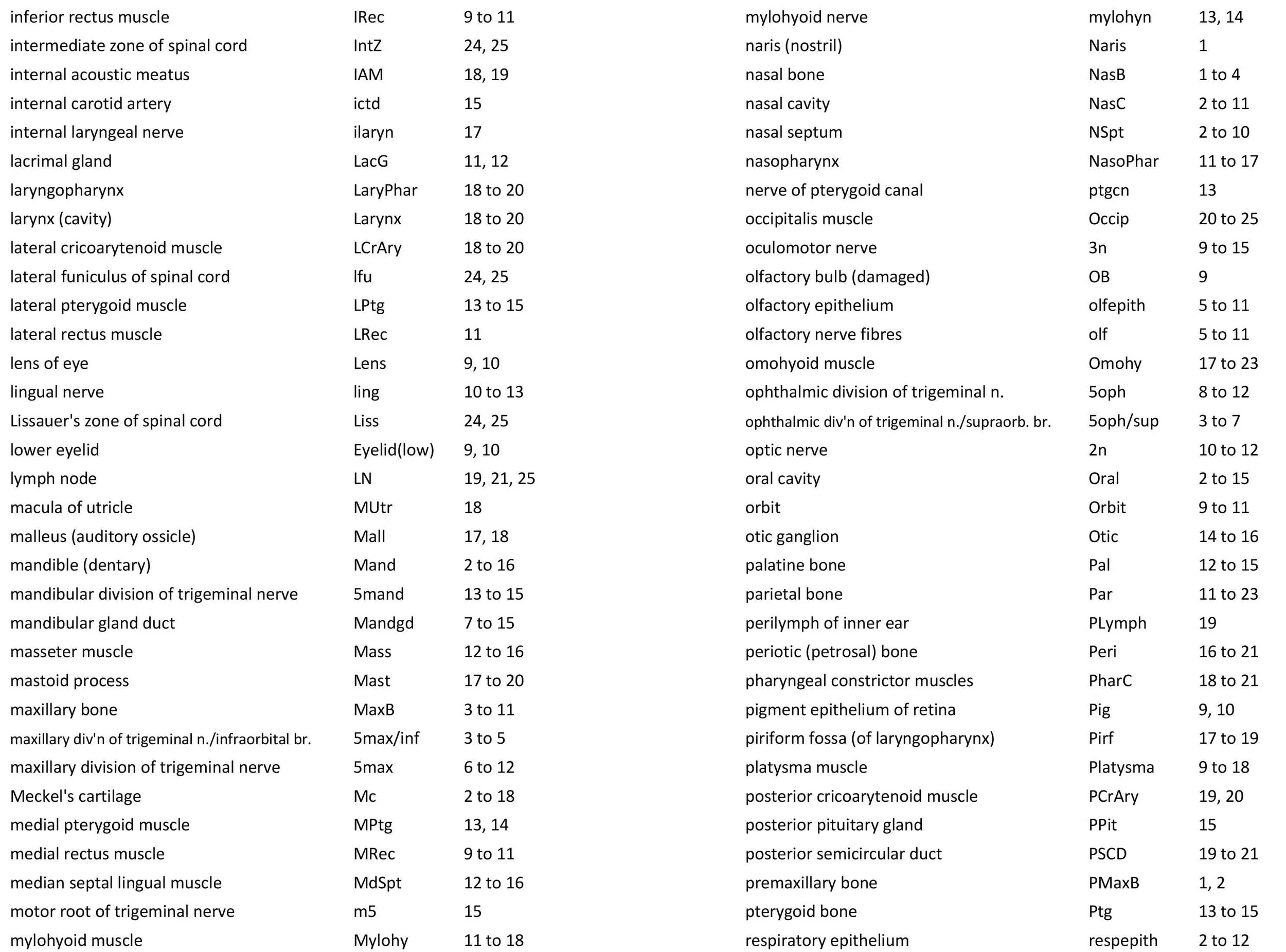
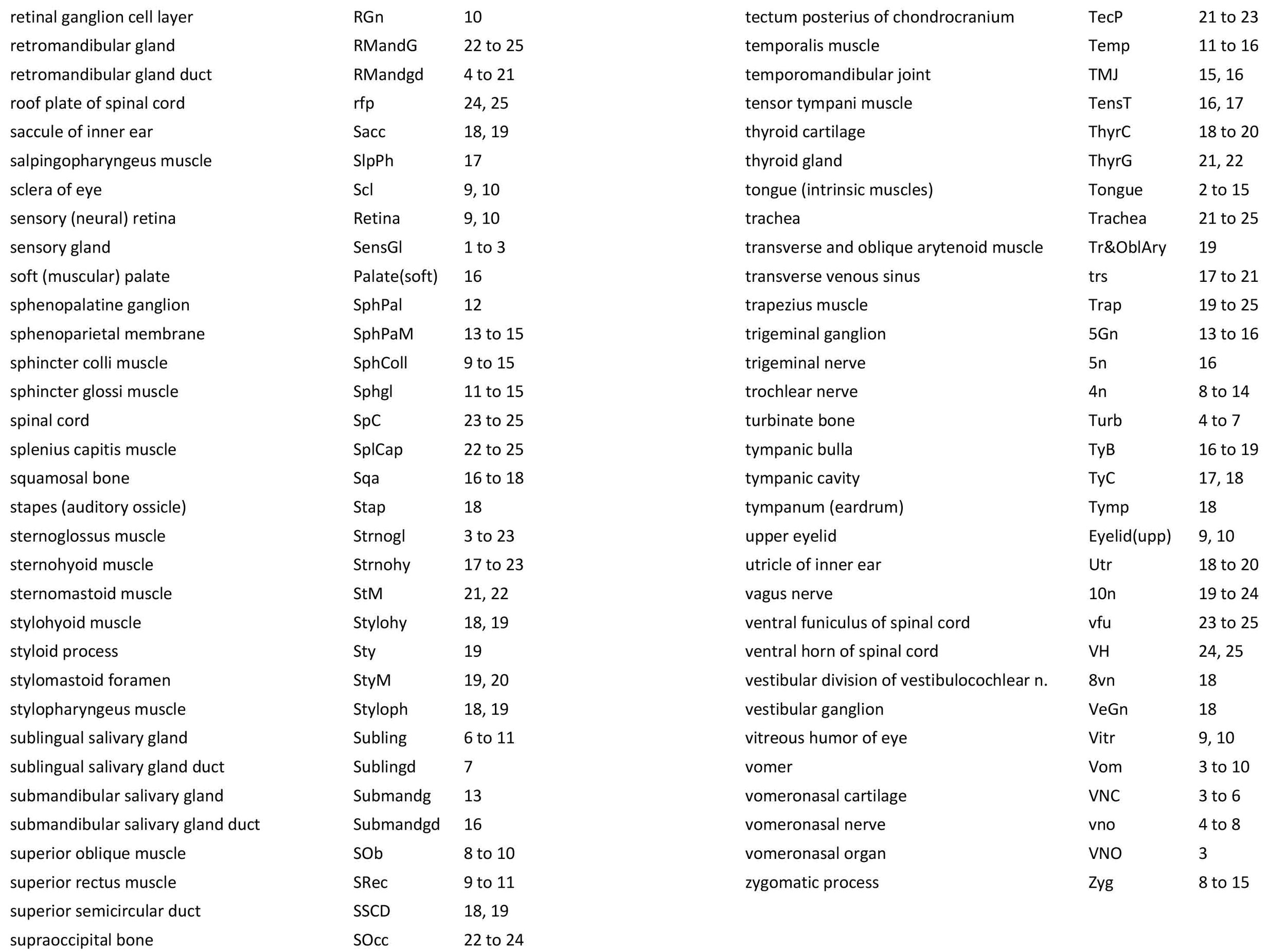
Atlas of the Head of a 24 mm GL (Hatchling) Echidna (M161)
Introduction
The short-beaked echidna specimen depicted here was labelled as “Echidna GL 24 mm”, where GL refers to greatest length. The subspecies is probably Tachyglossus aculeatus aculeatus, given that the most likely point of collection is eastern New South Wales near Sydney. An echidna puggle of 24 mm GL is probably 3 to 4 days after hatching (Ashwell, 2013)(where hatching occurs at about 14 to 16 mm GL) and the specimen would be stage 25 (Werneburg and Sánchez-Villagra, 2011).
Details on short-beaked echidna biology can be found at the Short-beaked Echidna page of this website.
Methods
The specimen illustrated here is M161 of the Hill collection stored at the Museum für Naturkunde in Berlin. The specimen had been collected in the late 19th century by JP Hill, embedded in paraffin wax and sectioned at 15 µm thickness. The stain was not identified on the specimen card, but may be a variant of carmine stain.
The sections at approximately 150 µm intervals were photographed with the aid of a Zeiss Axioplan2 fitted with an AxioCam MRc5 camera. All images were calibrated by photographing a scale bar at the same magnification and photomerged with Adobe Photoshop 2021. Merged images were placed in Adobe Illustrator 2021 and delineated.
Notes on the specimen
General observations
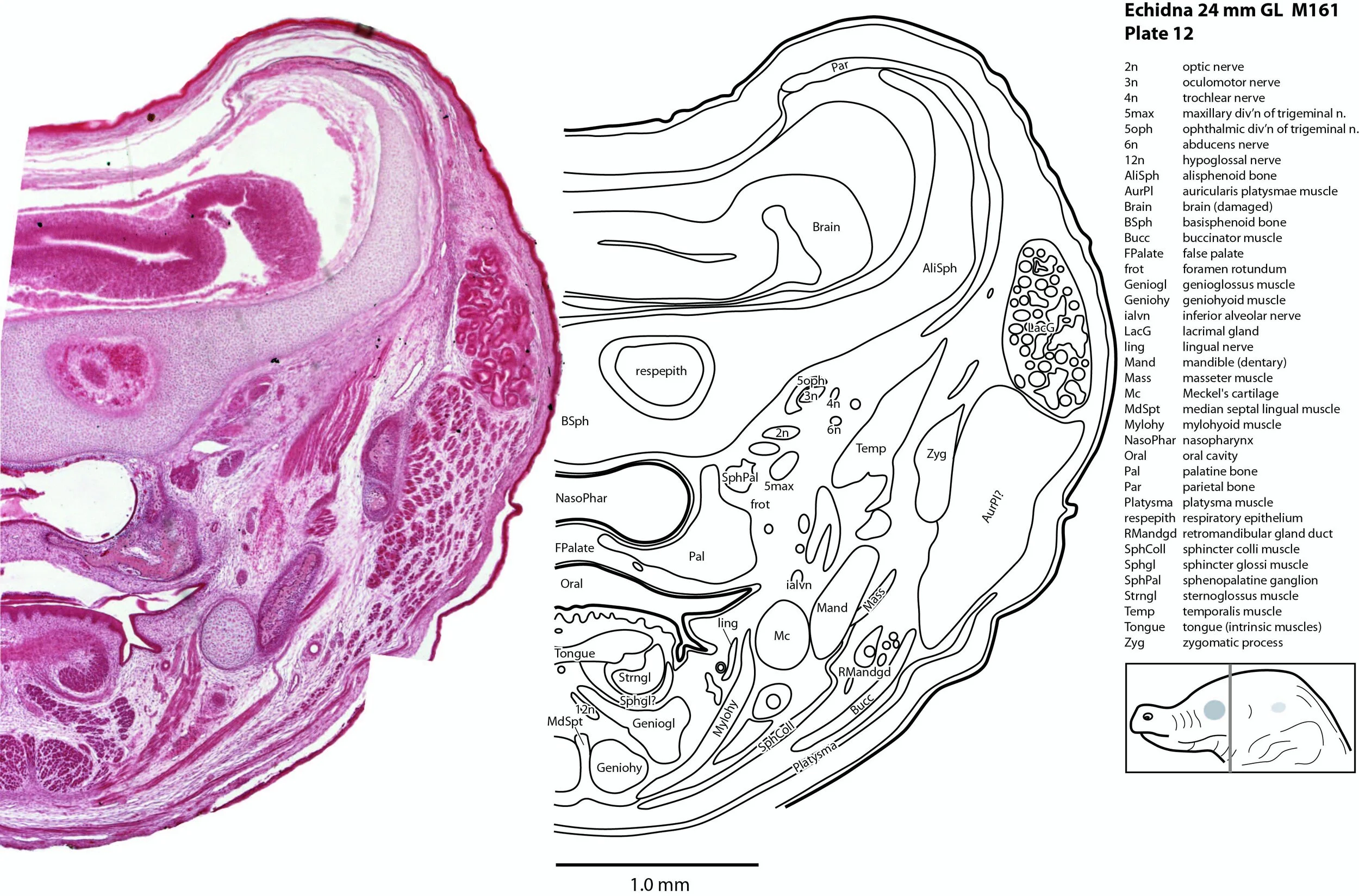
The brain is very poorly preserved, probably due to poor fixation of tissue within the cranium prior to embedding. It also appears that extraneous tissue (muscle, gut) has been stuffed into the cranial cavity prior to histological processing. Nevertheless, some intracranial structures (e.g. trigeminal ganglion, 5Gn – plates 13 to 16; pituitary gland, APit and PPit – plates 14, 15) and the spinal cord (SpC – plates 23 to 25) are present and much of the cranial peripheral nervous system is still visible.
Peripheral nervous system
The olfactory epithelium is extensive (olfepith – plates 5 to 11) and olfactory nerve fibres are visible (CN1, olf – plates 5 to 11), but the olfactory bulb is damaged. A vomeronasal organ VNO – plate 3) embedded within the vomeronasal cartilage (VNC – plates 3 to 6) can also be seen, and a vomeronasal nerve (vno – plates 4 to 8) traced towards the cranial cavity.
A nascent optic nerve (2n – plates 10 to 12) is attached to the eye bulb (see below) and can be traced some distance through the orbit, but its connection with the brain is lost.
The origins of the eye muscle nerves (3n, 4n, 6n) have been lost, but these nerves can be seen within the orbit (3n – plates 9 to 15; 4n – plates 8 to 15; 6n – plates 11 to 16) and adjacent to the respective eye muscles they supply.
The trigeminal ganglion is large (5Gn – plates 13 to 16) and the stub of the trigeminal nerve (CN5) is present but friable (5n – plate 16). The motor root (m5 – plate 15) can be seen passing through the trigeminal ganglion. All three divisions of the trigeminal nerve can be seen (5oph – plates 8 to 12; 5max – plates 6 to 12; 5mand – plates 13 to 15), as well as several major branches such as the supraorbital branch (5oph/sup – plates 3 to 7), infraorbital branch (5max/inf – plates 3 to 5), and inferior alveolar nerve (ialvn – plates 3 to 12).
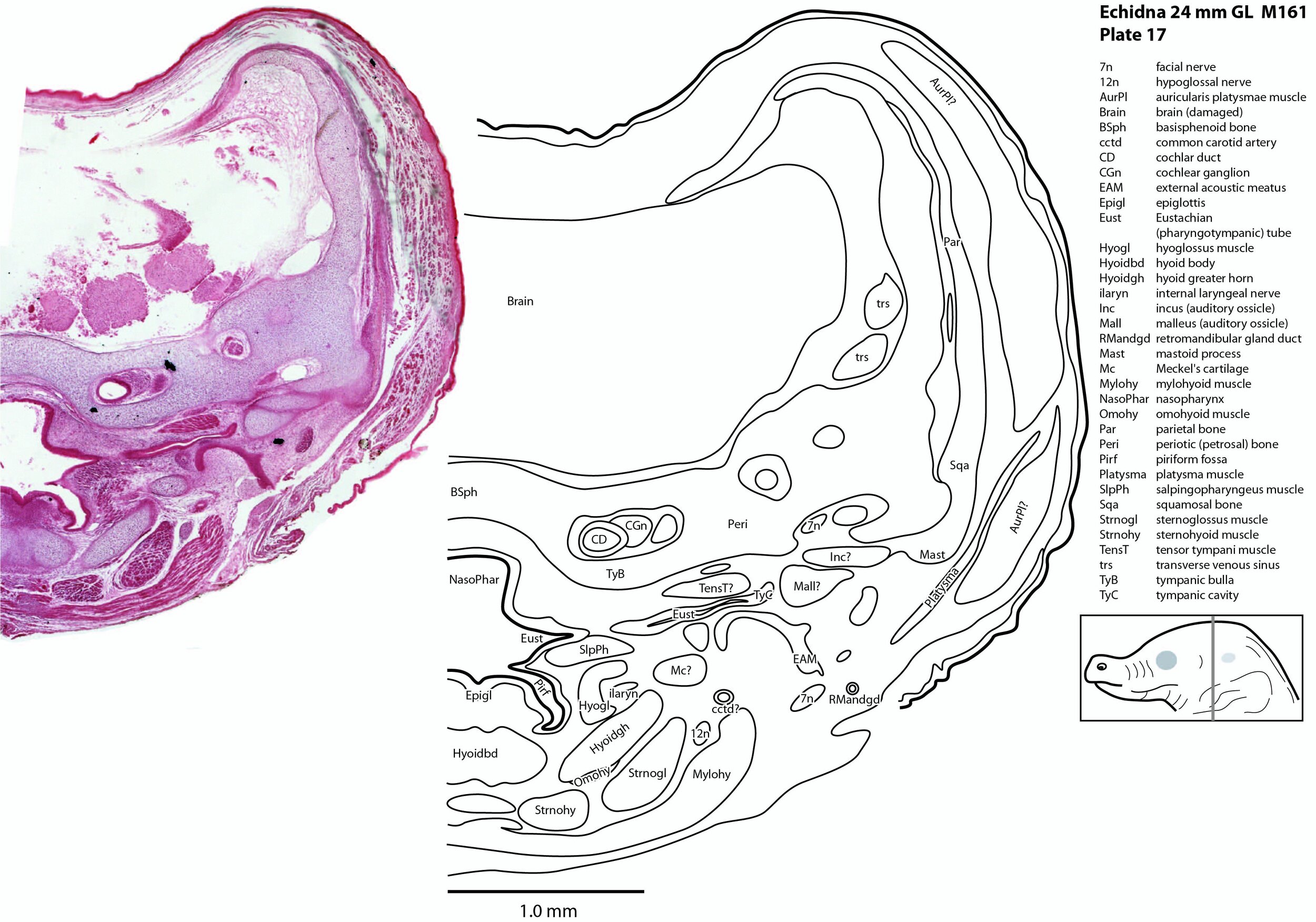
The facial nerve is large (plates 17 to 20) and visible exiting the stylomastoid foramen (StyM - plates 19 and 20). Its large size may be due to its role in supplying the large volume of superficial platysmae musculature (please see below).
Central auditory structures have been damaged, but a stub of the vestibular division of the vestibulocochlear nerve is visible (8vn – plate 18) and both cochlear and vestibular ganglia are still intact CGn – plates 17 to 19; VeGn – plate 18).
The glossopharyngeal nerve (9n – plates 18 to 21) is best seen close to the muscle it supplies (stylopharyngeus).
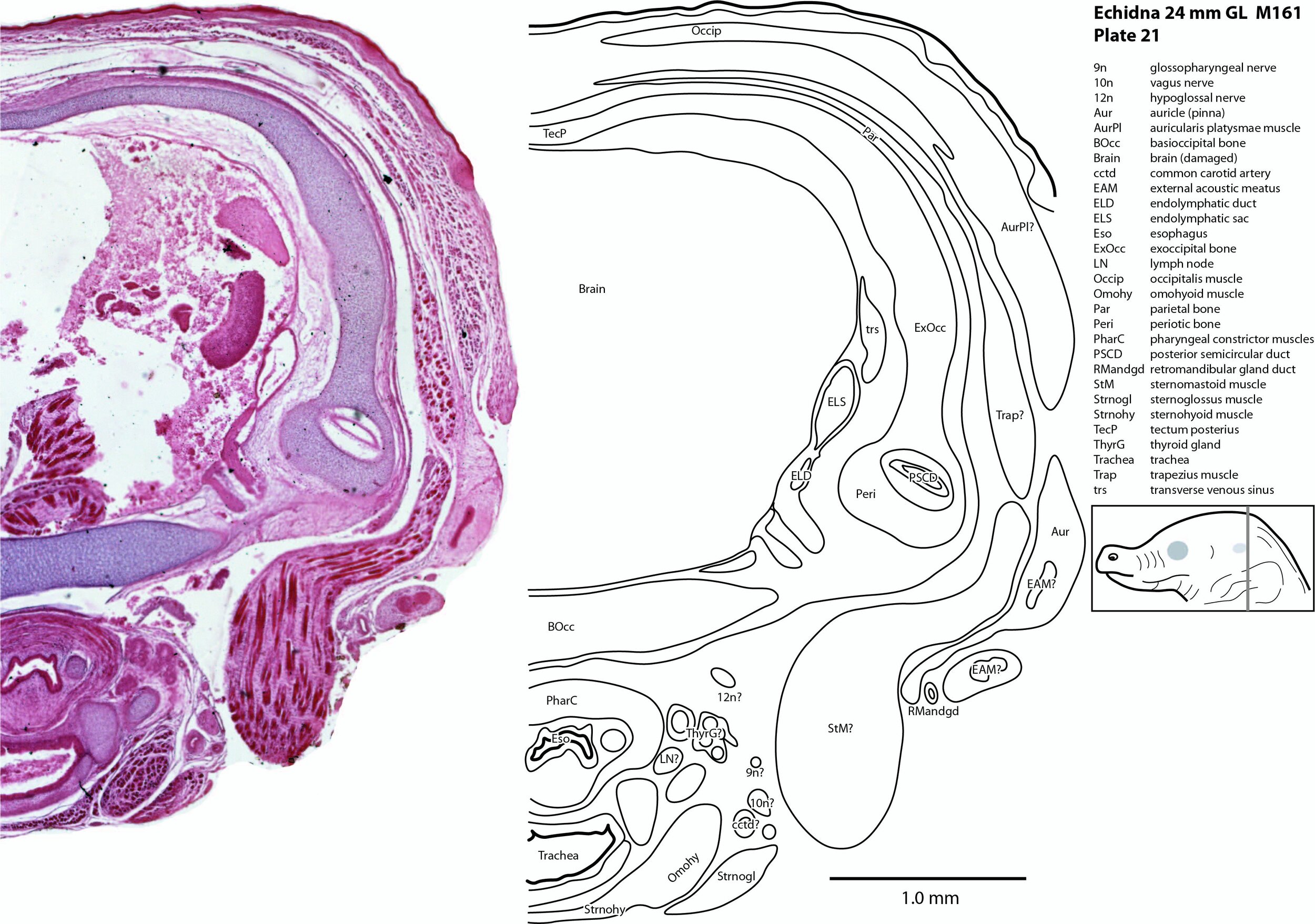
The vagus nerve (10n) is best seen adjacent to the airway and esophagus (plates 19 to 24), whereas the large hypoglossal nerve (12n) has a long course (plates 12 to 21) supplying the large volume of lingual musculature.
Sense organs
The Anlagen of (electro)sensory glands are visible in the beak (SnsGl – plates 1 to 3), but are too immature to be functional.
The olfactory epithelium is extensive in area, but thin (olfepith – plates 5 to 11).
The eye is still very rudimentary, with only a very thin retinal ganglion cell layer (RGn – plate 10) present. The eye is completely covered by skin (plates 9, 10) and the palpebral fissure closed, but the lens and cornea are present.
The ear is also rudimentary. The external acoustic meatus is closed (EAM – plates 17 to 25), the tympanic cavity a narrow slit (TyC – plates 17, 18) and the auditory ossicles represented only by cartilaginous models (see malleus, incus, and stapes – plates 17, 18). Maculae (sensory zones) of the organ of Corti and vestibular apparatus are only rudimentary.
Musculoskeletal system
Most of the skull is a cartilaginous model (chondrocranium) at this stage, with some neuro- and viscerocranial bones forming within membranes around the cartilage.
Rather than use transient chondrocranial nomenclature, labels for the adult bone that would develop from the cartilage model have been assigned. The sole exception to this is the tectum posterius (TecP – plates 21 to 23), a caudal midline roof feature of the chondrocranium (Griffiths, 1978).
Nomenclature for musculature is mainly based on Griffiths (1978). The two most striking muscle groups are the superficial (subdermal) facial group and the lingual musculature.
The superficial facial group includes a frontalis platysmae (FroPl – plates 5 to 10), a (pre)auricularis platysmae (AurPl – plates 8 to 23), and an occipitalis muscle (Occip – plates 20 to 25). Presumably these muscles are important for shifting the spines forward to protect the head during the defensive posture.
The lingual muscles include the sternoglossus (Strnogl – plates 3 to 23) and median septal (MdSpt – plates 12 to 16) muscles (both described by Griffiths, 1978), the first of which must be a powerful lingual retractor, a genioglossus (Geniogl – plates 3 to 14) presumably a protrusor of the tongue, as well as a muscle which spirals obliquely around the distal sternoglossus, and which has been called here sphincter glossi (Sphgl – plates 11 to 15) because it would be able to compress the sternoglossal belly and cause protrusion. A small hyoglossus (Hyogl – plates 14 to 17), presumably a lingual retractor and depressor, was also found.
The masticatory muscles are relatively small in volume, especially the masseter (Mass – plates 12 to 16) and temporalis (Temp – plates 11 to 16), consistent with the limited load requirement for crushing of food. Some protraction and lateral movement of the mandible (dentary) would be needed for crushing arthropods, and these are provided by the lateral and medial pterygoid muscles (LPtg – plates 13 to 15; MPtg – plates 13, 14).
Acknowledgements
I would like to thank Dr Peter Giere of the MfN, Berlin Germany, for access to the collection and for all his kind help during the work.
References
Ashwell KW (2013) Embryology and post-hatching development of the monotremes. In KWS Ashwell (Ed.), Neurobiology of Monotremes: Brain Evolution in Our Distant Mammalian Cousins (1st ed., pp. 31-46). CSIRO.
Griffiths M (1978) The Biology of the Monotremes. Academic, New York.
Werneburg I and Sánchez-Villagra MR (2011) The early development of the echidna, Tachyglossus aculeatus(Mammalia: Monotremata), and patterns of mammalian development. Acta Zoologica (Stockholm) 92, 75–88.