Atlas of the Brain of a 7 week Human Embryo, Carnegie #462
Introduction
This specimen is on the embryo/fetus cusp, meaning that all major organs have been formed, but considerable cytogenesis (cell generation) and differentiation must still take place. In the case of the brain, this means generation (neurogenesis) of most of the 86 thousand million neurons required for an adult human brain and their accompanying 80 to 90 thousand million glia. Most of the neurons present at 7 weeks are those large body/long axon macroneurons in the brainstem that will form the long tracts of the adult brain. Microneurons of the brainstem and all neurons large and small of the cerebral and cerebellar cortices are yet to be produced.
Methods
The specimen illustrated here is Carnegie #462, held at the National Museum for Health and Medicine in Silver Springs, Maryland. On the virtual human embryo website (virtualhumanembryo.lsuhsc.edu) the specimen is said to be stage 20, GL of 20 mm post-fixation. Post-dehydration length calculated from the product of section count and thickness (i.e. 397 x 0.04 mm) is 15.9 mm. Estimated post-conception age is variously given as 49 days, or 51 to 53 days.
The specimen is said to have been acquired in 1910, fixed in formol, embedded in paraffin, sectioned transversely to the long axis of the embryo at a thickness of 40 µm, and the sections stained with alum cochineal (carmine).
Raw images were collected by Ken Ashwell at the NMHM in 2014, using a Nikon E800 microscope with a Ludl Biopoint automated stage and a MAC 2000 controller. Photomicrographs were taken with Micropublisher V5, with ImagePro Plus software from Media Cybernetics.
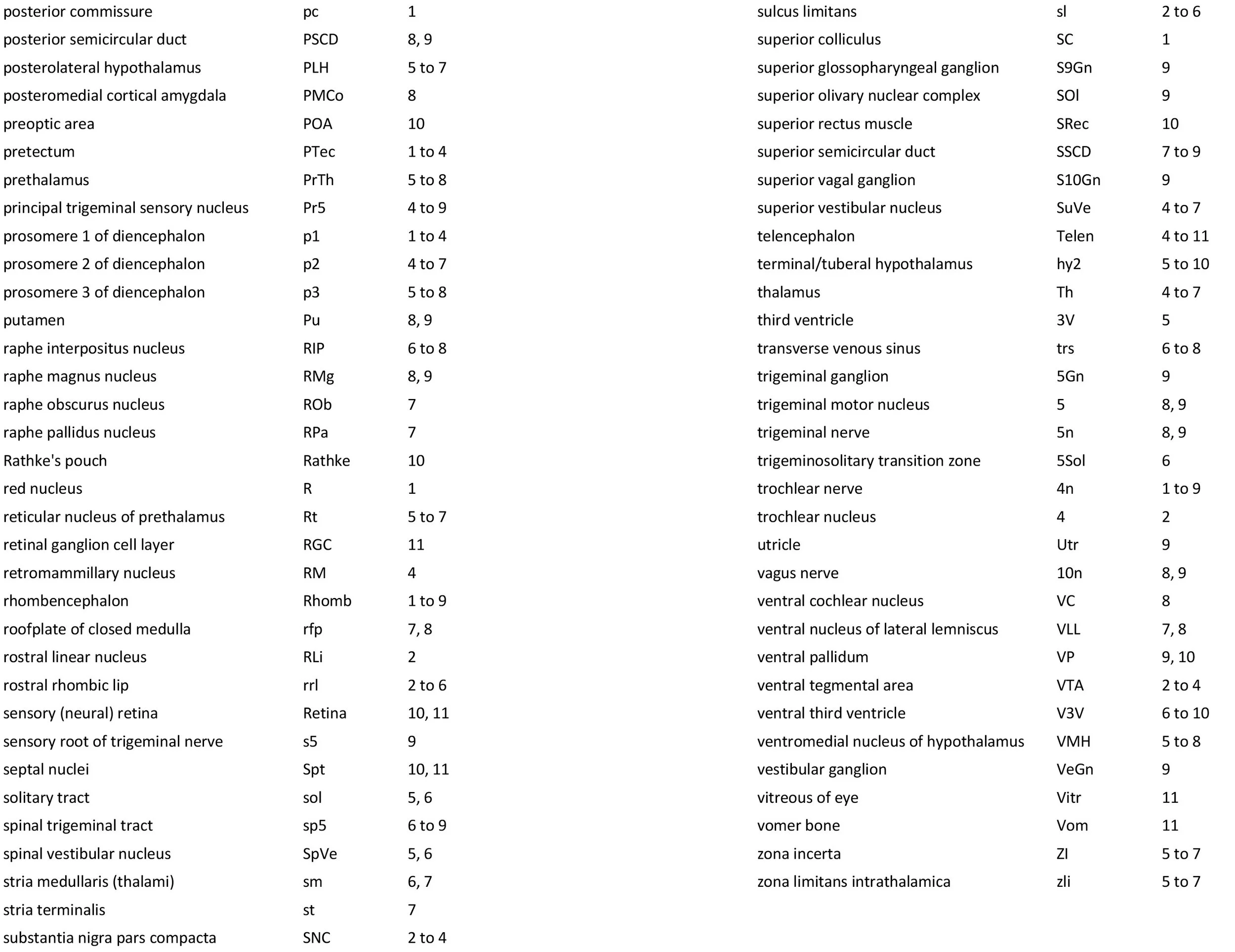
All images were calibrated by photographing a scale bar at the same magnification. Images were photomontaged and optimised for contrast using Adobe Photoshop 2023 before being placed in Adobe Illustrator 2023 for delineation and labelling. Developmental regions (i.e. neuroepithelium) destined to give rise to adult structures have been denoted by the adult structure’s name with an asterisk (e.g. Cx* denotes the developmental field of the cerebral isocortex).
A line diagram of a stage 20 human embryo has been used for the finder illustration indicating section position.
Notes on the specimen
General observations
The most differentiated part of the brain is the rhombencephalon (brainstem) where definitive nuclear groups have begun to appear. The forebrain shows prosomeric divisions of the neural tube, but few post-mitotic neurons. The cerebral isocortex is rudimentary and mostly consists of proliferative populations. The cerebellum is also rudimentary and consists of only the cerebellar primordium of the rostral rhombic lip.
Central nervous system
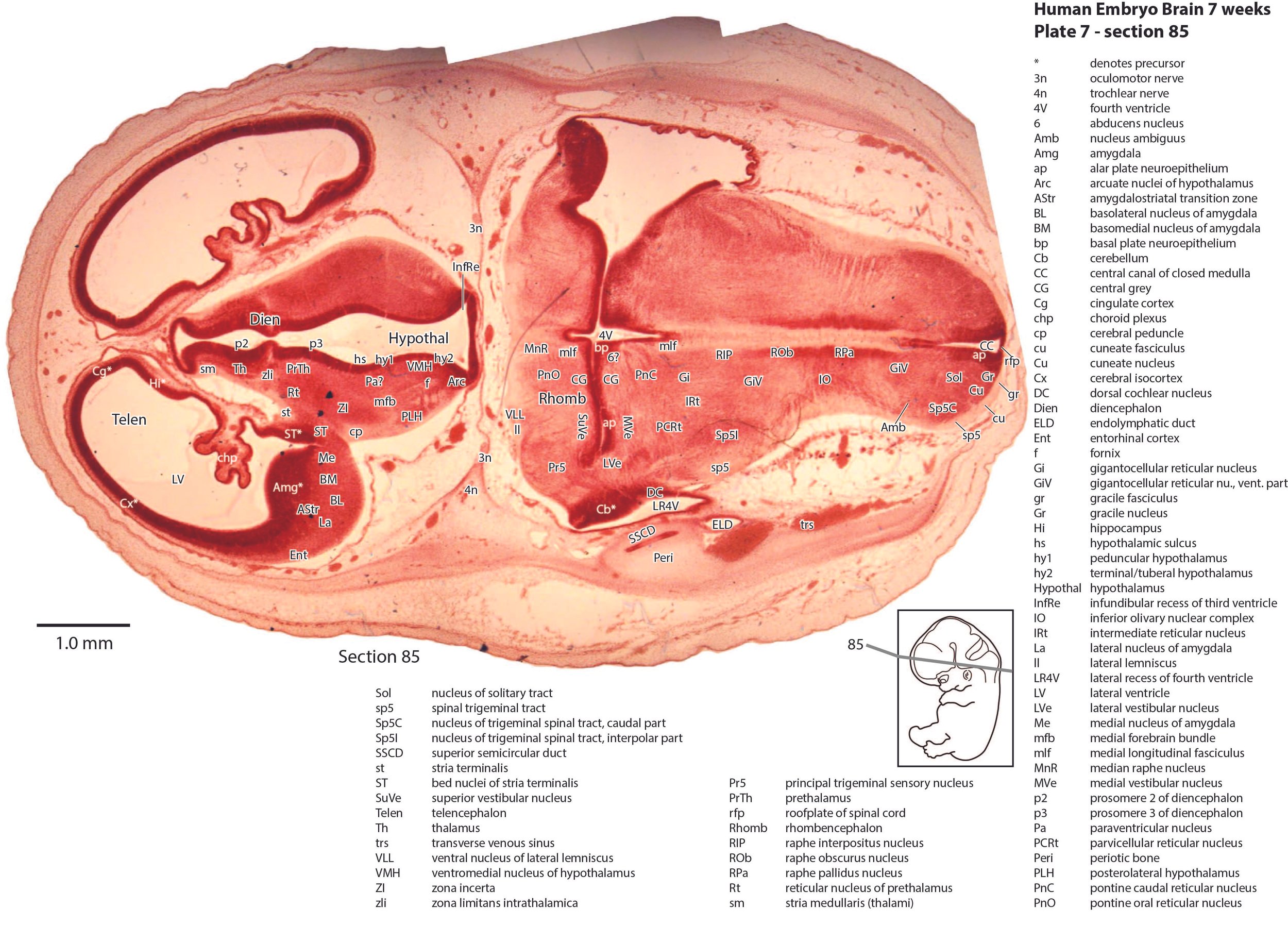
The olfactory bulb (plate 11) is still rudimentary, with a very thick generative (mitotic) neuroepithelium and very few postmitotic neurons released. The telencephalon has a thin pallium (Cx, Cg, Ent), with a thin external preplate (plates 4 to 11) over the dorsal and medial pallium. The hippocampus (Hi in plates 5 to 10) is also rudimentary, represented by a curved region of pallium projecting into the lateral ventricle. It has a thick proliferative zone, surmounted by a thin preplate zone, which is mostly fibrous with very few neurons. The subpallial telencephalon has the usual lateral ganglionic eminence (lge in plates 8 to 11) and medial ganglionic eminence (mge in plate 9, 10). There are some early-generated striatal and pallidal neurons (Cd in plates 8 to 11, Pu in plates 8 and 9, GP in plates 8 to 10, VP in plates 9 and 10), but it is difficult to distinguish internal structure among those neuronal populations. Only the notional positions of striatal and pallidal components have been labelled.
The hypothalamus is divided into peduncular (hy1), tuberal/terminal (hy2), and acroterminal (hyat) proliferative components (variously in plates 5 to 10), surrounded by postmitotic populations of neurons in arcuate (Arc in plates 6 to 10), paraventricular (Pa in plates 6 to 8), and ventromedial (VMH in plates 5 to 8) nuclear groups, as well as lateral (LH in plates 8 and 9) and posterolateral neuronal populations (PLH in plates 5 to 7).
The diencephalon has the typical prosomeric structure. Prosomeres 1 (pretectum), 2 (dorsal thalamus), and 3 (prethalamus) can be identified as proliferative zones (see plates 1 to 8), but postmitotic populations are very small, particularly in the dorsal thalamus (Th in plates 4 to 7).
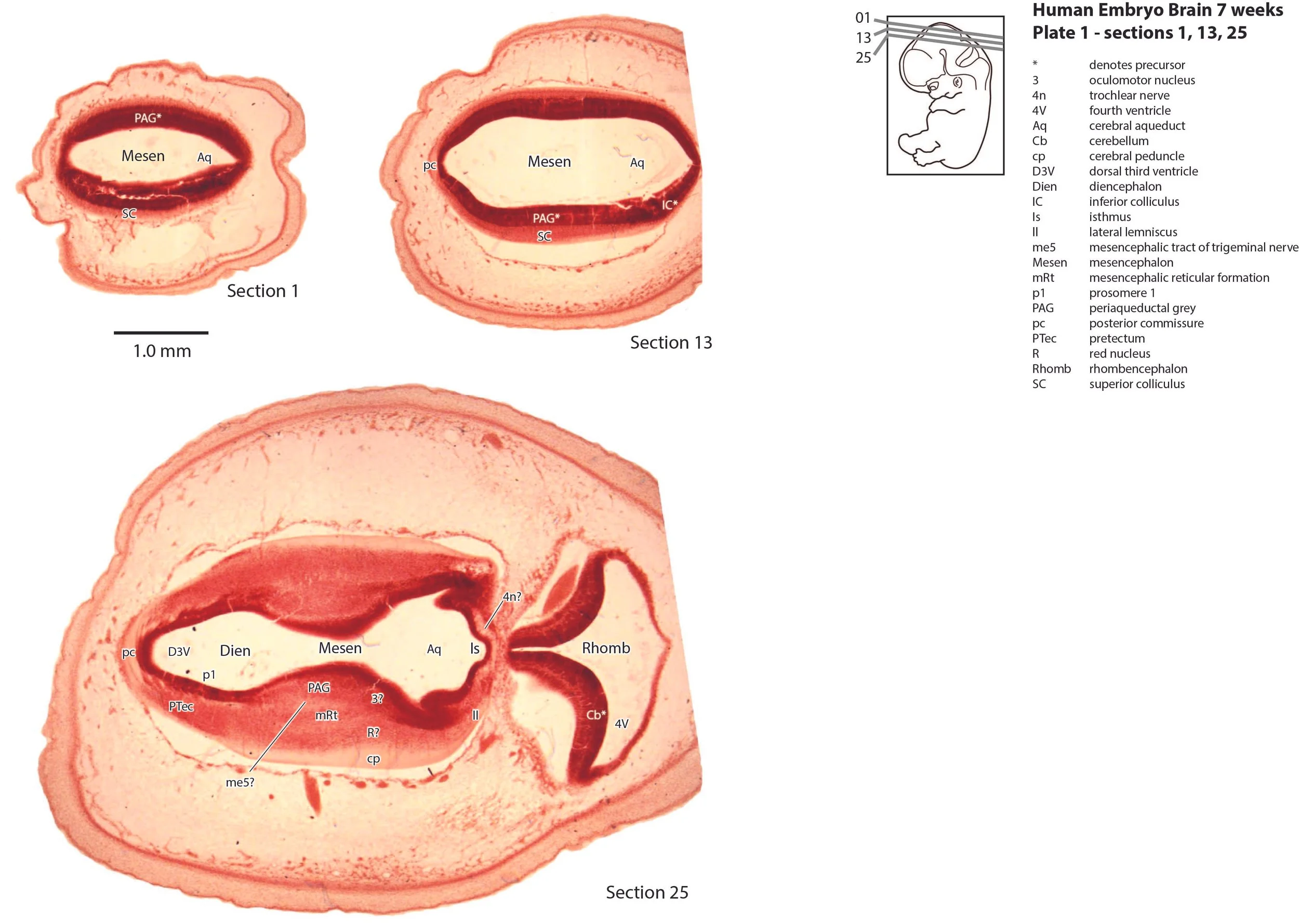
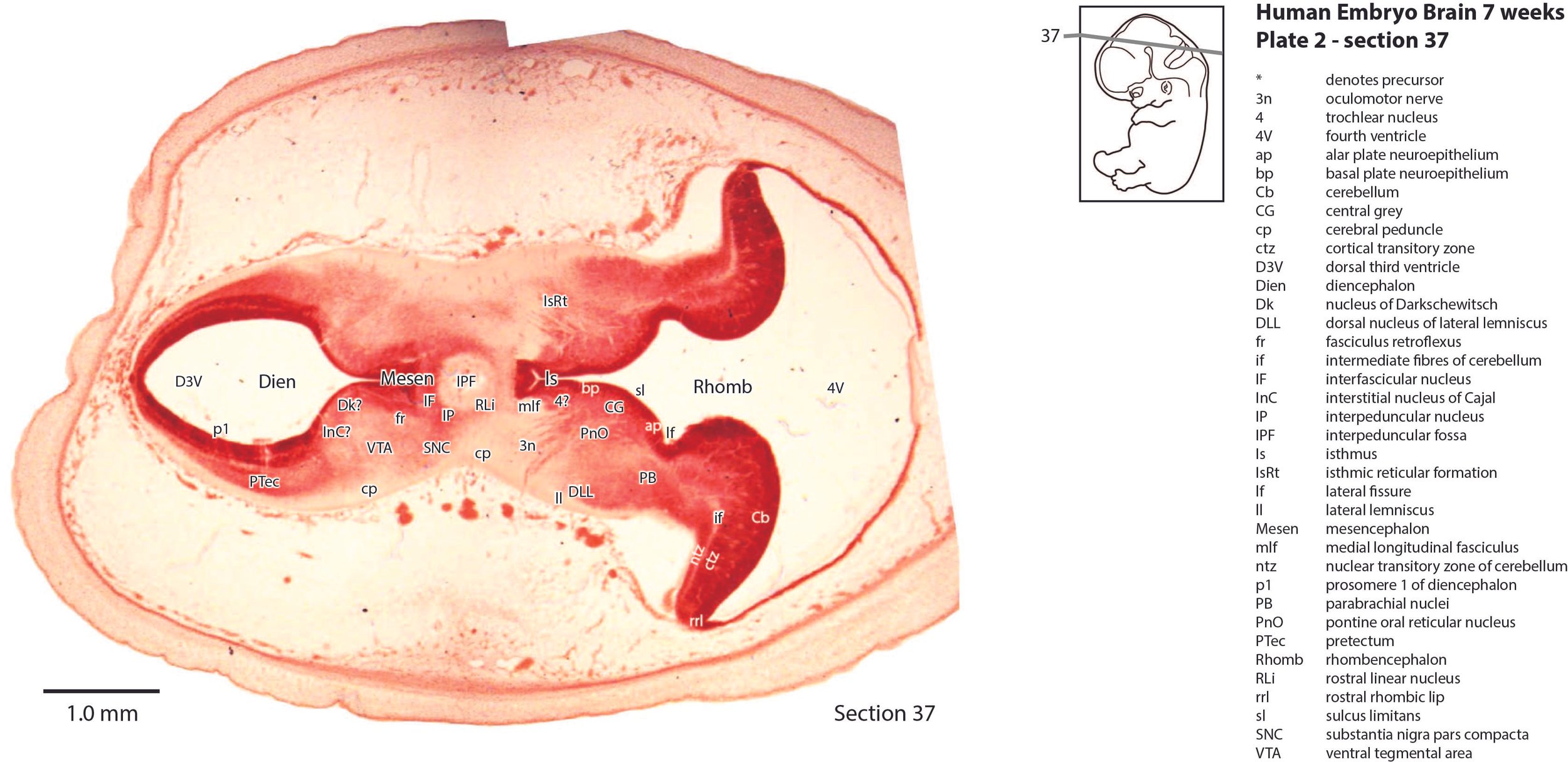
The mesencephalon has a rudimentary superior colliculus (SC in plate 1) and inferior colliculus (IC in plate 1), where most of the neuraxis wall is made up of the proliferative neuroepithelium, and very few postmitotic neurons. The mesencephalic tegmentum is poorly developed. Only the notional position of major nuclei can be marked, because differentiation of nuclei and fibre tracts is poor.
The cerebellum (plates 1 to 7) is a lid extending over the fourth ventricle and rimmed by the rostral rhombic lip (rrl in plates 2 to 6). No cortical or deep nuclear differentiation is evident.
The brainstem is the most differentiated part of the brain. The trigeminal sensory nuclear complex (Pr5, Sp5O, Sp5I, Sp5C; variously in plates 5 to 9) occupies most of the lateral sensory column of the brainstem. As in other mammals, the medial magnocellular reticular formation (Gi, GiV, LPGi; plates 6 to 8) is broader that the parvicellular reticular formation (PCRt; plates 6 to 8). A putative lateral vestibulospinal tract (lvs in plates 5, 6) suggests an early projection from the vestibular nuclear complex to the spinal cord. The dorsal column pathways (gr and cu in plates 6 to 8) are still very thin, indicating that ascending sensory pathways are rudimentary.
Peripheral nervous system
Most cranial nerves can be identified. Cranial nerve 1 is multiple fibres (olf in plate 11) running from the olfactory epithelium to the nascent olfactory bulb. Cranial nerve 2 (optic) is not developed, but an optic stalk (os in plate 10) extends from the hypothalamus to the eye cup. The eye muscle nerves 3n and 4n can be traced from the brainstem to the orbit (3n in plates 2 to 10, 4n in plates 1 to 9). 6n is not visible. The trigeminal nerve (5n in plates 8, 9) runs from the trigeminal ganglion (5Gn in plate 9) to the brainstem. The facial nerve can be traced from the facial motor nucleus (7 in plates 8 and 9) through the geniculate ganglion (Gen in plate9) and externally as 7n (plate 9). Components of the vestibulocochlear nerve pathway include the vestibular ganglion (VeGn in plate 9) and cochlear nuclei (DC and VC in plates 9 and 8, respectively). The emergent glossopharyngeal/vagal/accessory nerve rootlets are visible as a group, but only the vagus nerve (10n in plates 8 and 9), superior glossopharyngeal ganglion (S9Gn in plate 9), superior vagal ganglion (S10Gn in plate 9) can be picked out clearly. The hypoglossal nerve is below the plane of the last plate, but the hypoglossal nucleus is visible (12 in plates 5 and 6)
Acknowledgements
I would like to thank Dr Elizabeth Lockett of the NMHM for access to the collection and for all her kind help during the work.