A Cytoarchitectonic Atlas of the Brain of the Common Bottlenose or Atlantic Dolphin (Tursiops truncatus)
Introduction
The common bottlenose or Atlantic dolphin has an adult length of 2.0 to 3.8 m and adult weight from 220 to over 500 kg, with males larger than females. It is found in temperate and tropical oceans throughout the world. Longevity is estimated at 40 to 50 years.
From 1998, Tursiops aduncus (see whole dolphin brains from the South Australian Museum) has been considered a separate species within the genus, but the taxonomic status of T. aduncus may change.
Brain size of adult Tursiops truncatus is about 1.6 kg, approximately 16% larger than humans.
Materials and Methods
The brain depicted here is from the Yakovlev collection and the Nissl-stained sections are held at the National Museum for Health and Medicine in Silver Springs, Maryland, USA. Selected horizontal plane sections in a regular sequence (please see section numbers on plates) were scanned with the aid of an Epson 11000.
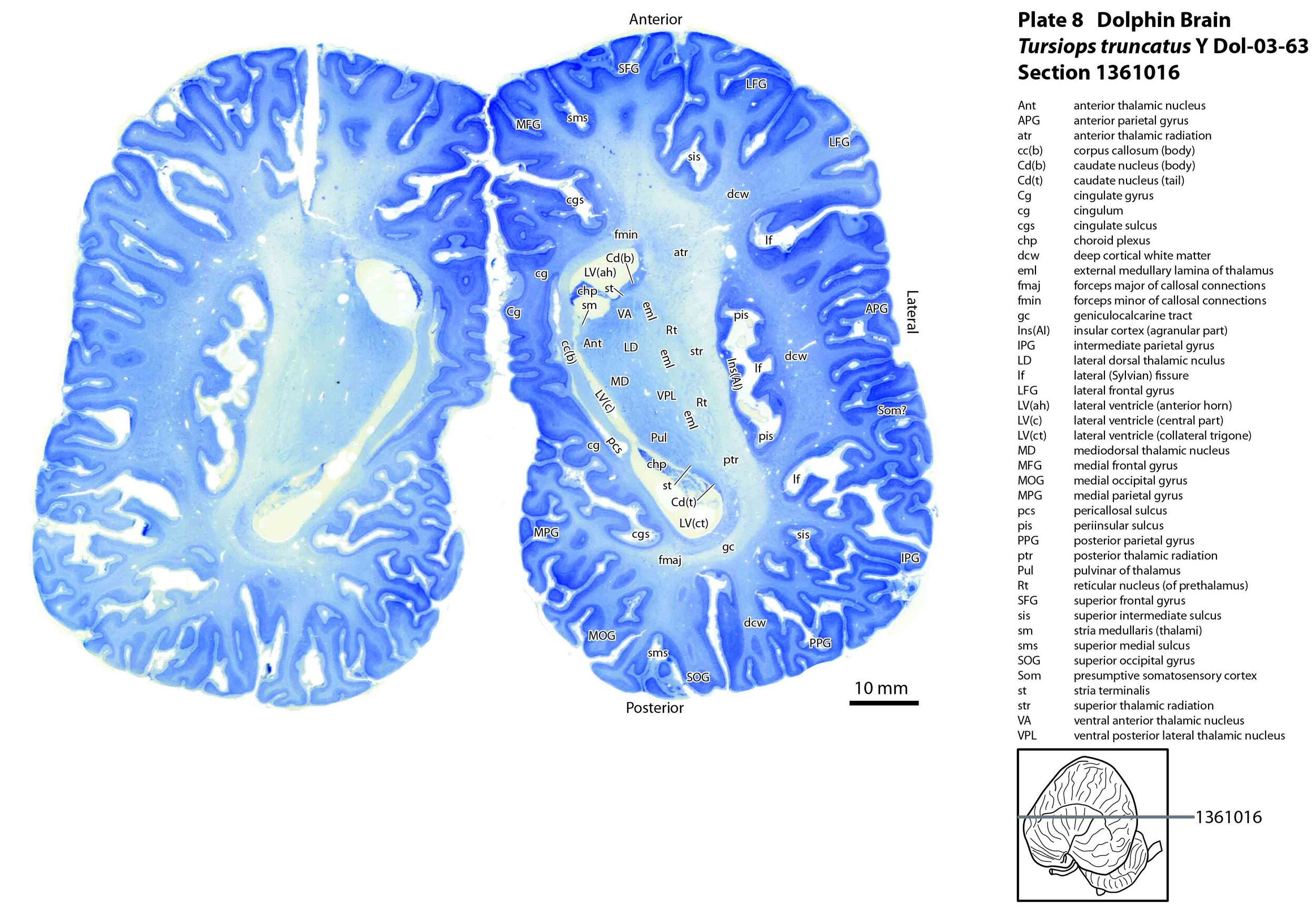
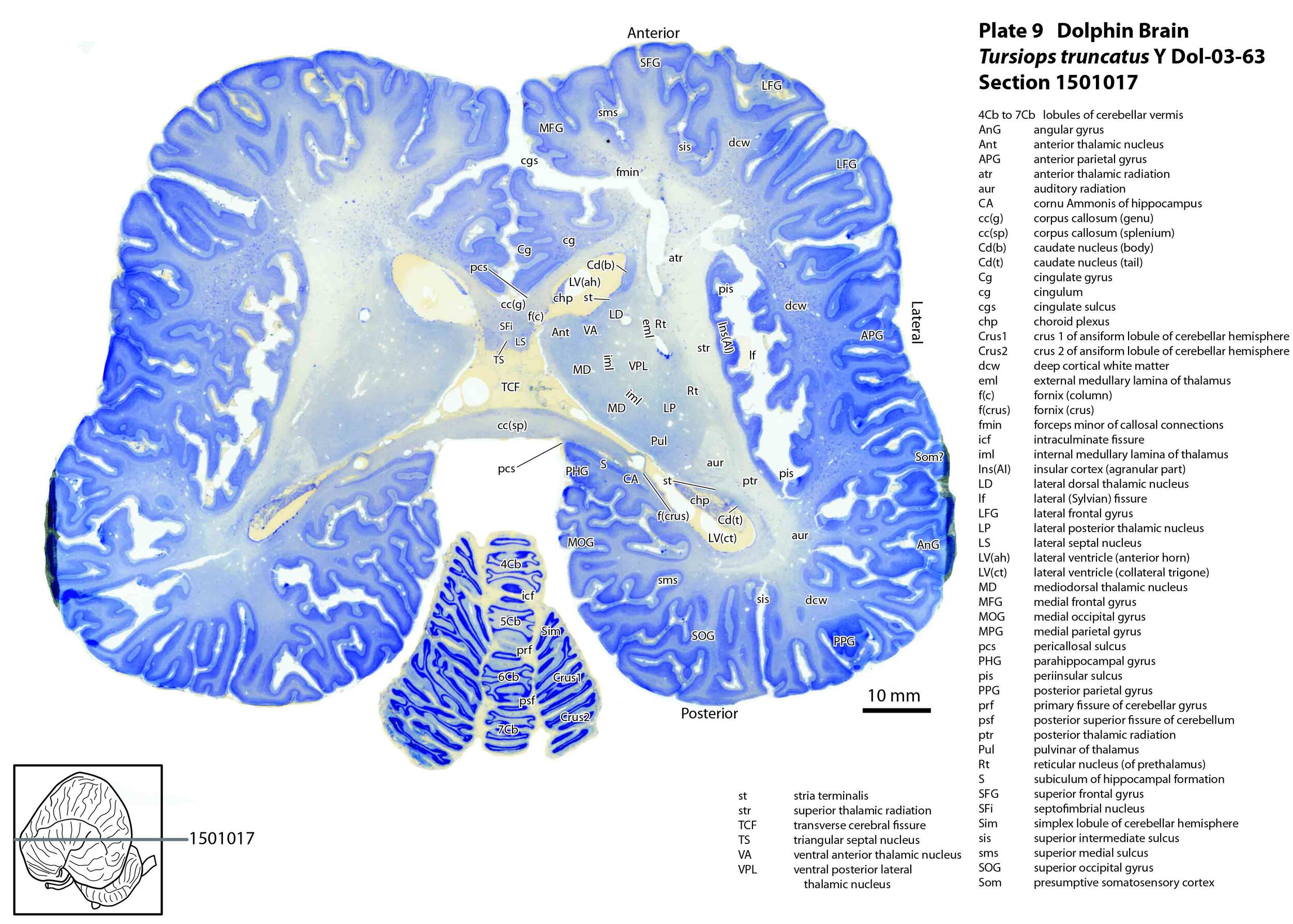
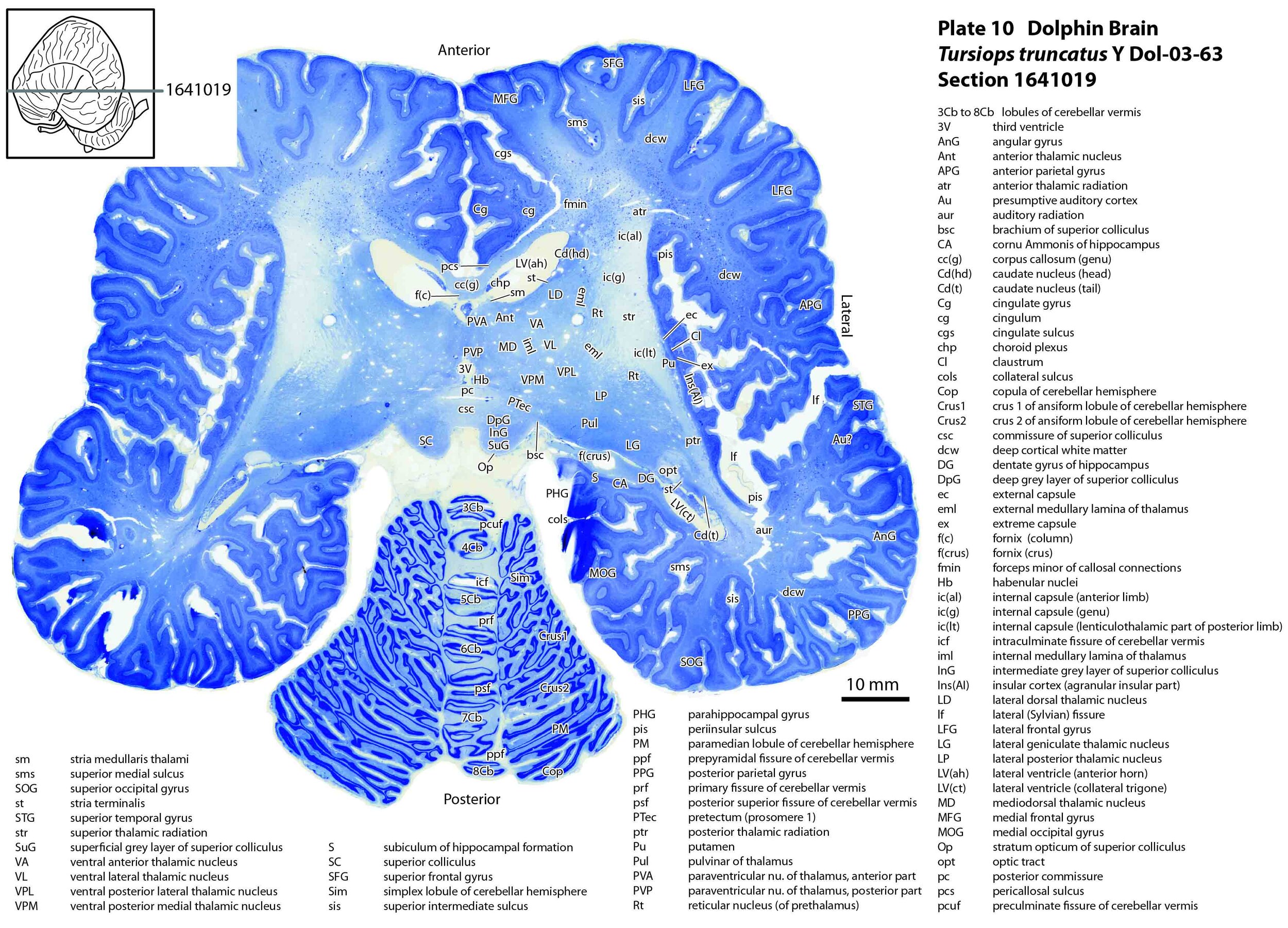
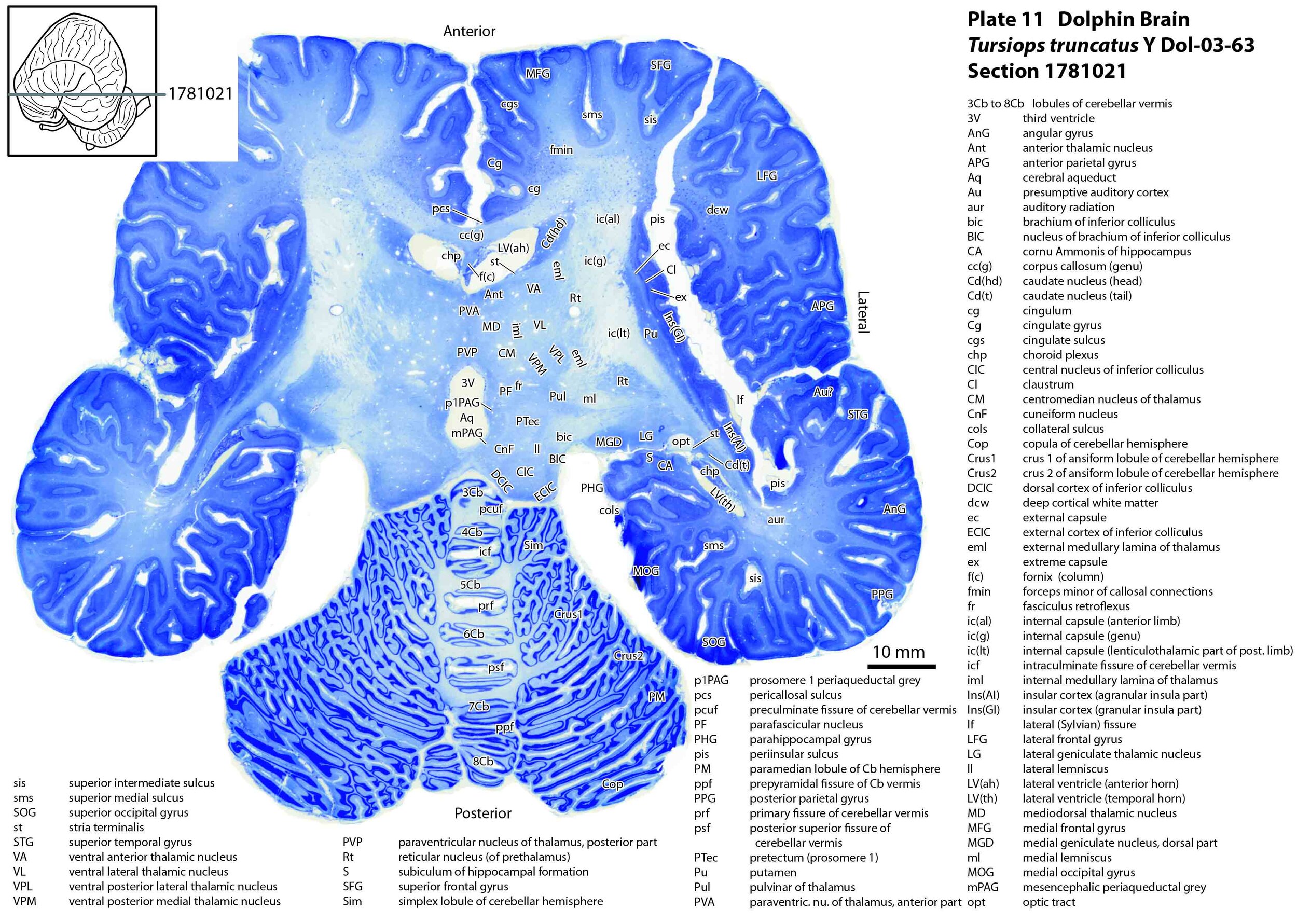
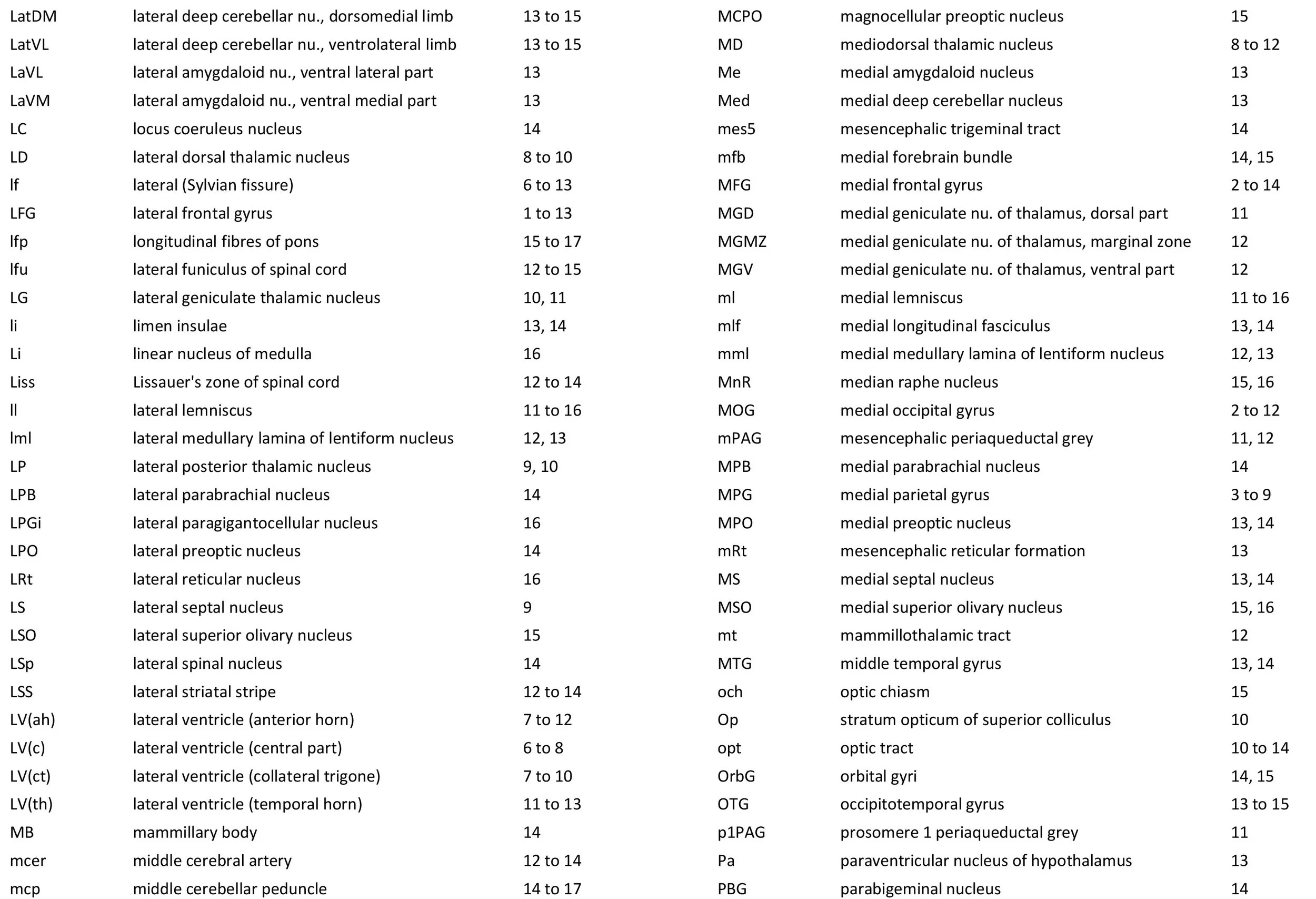
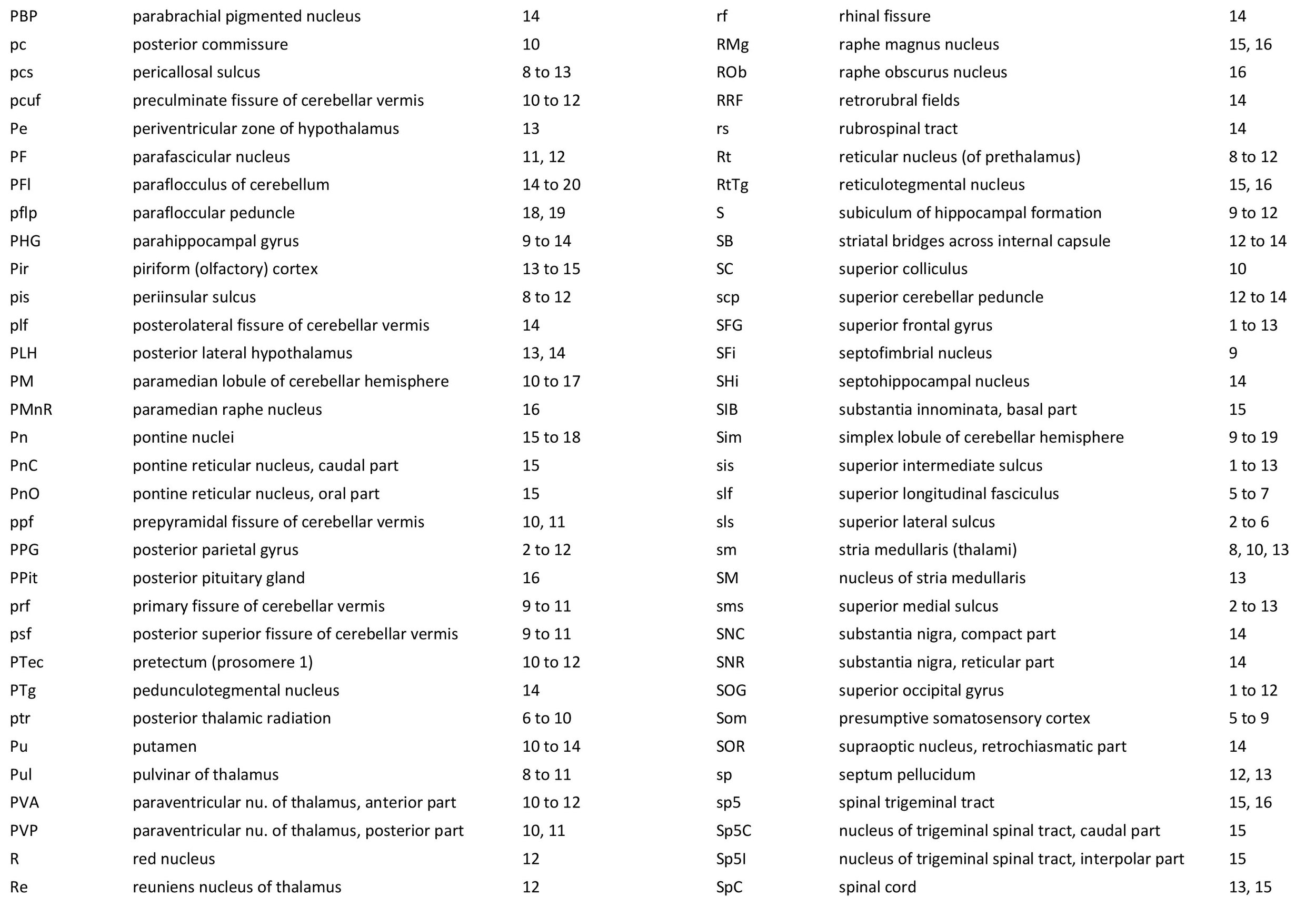
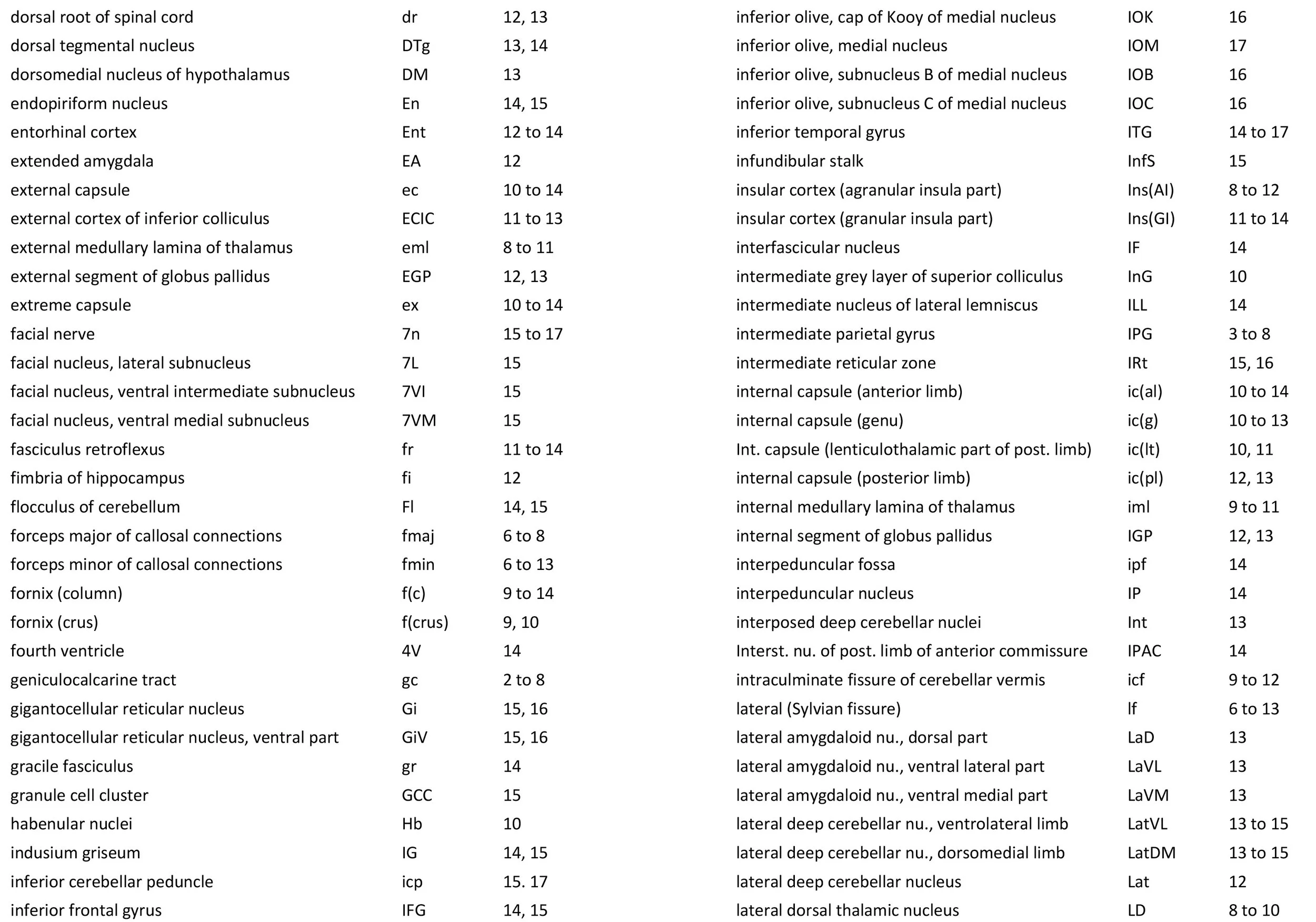
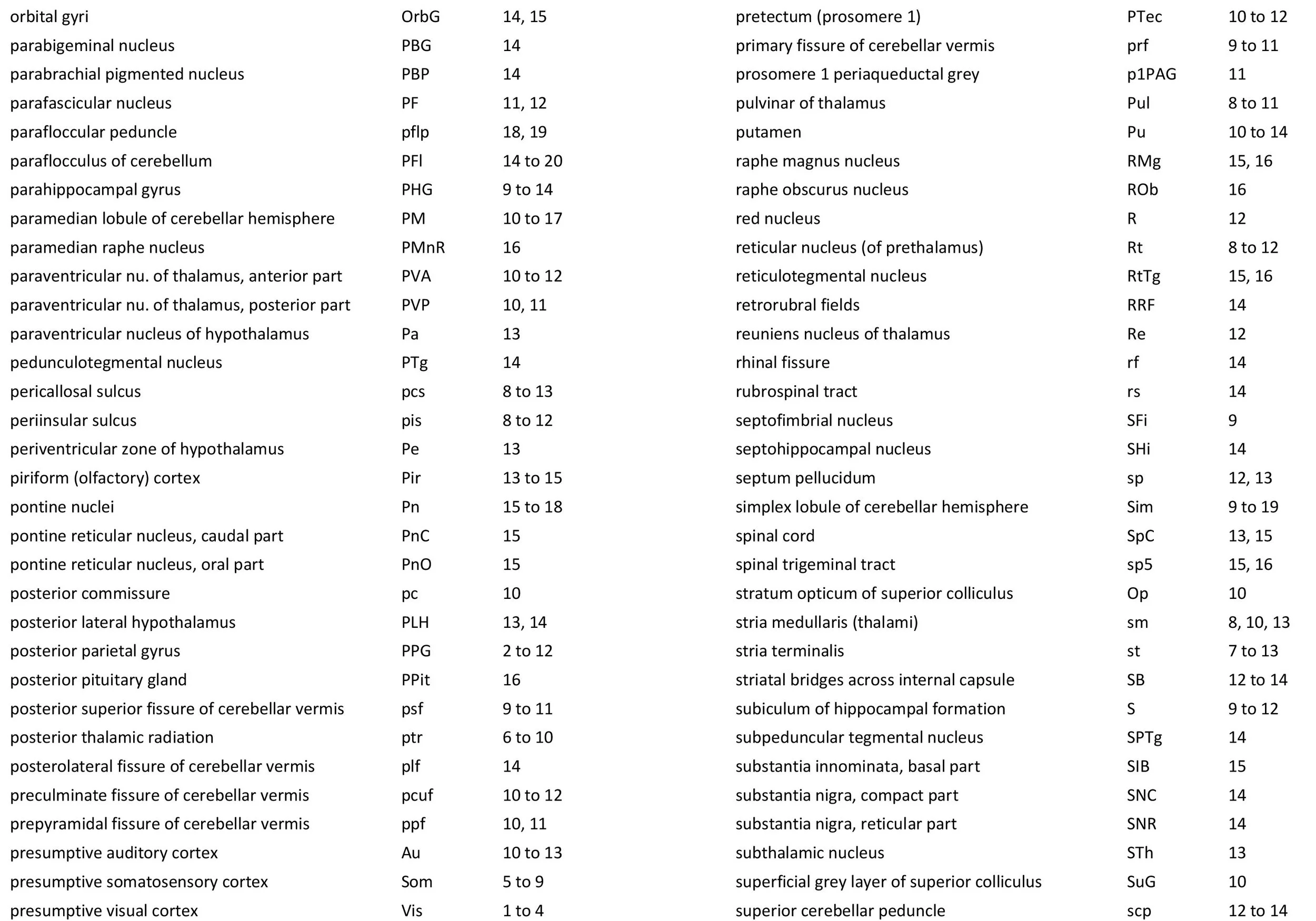
The images were cropped and adjusted for optimal contrast levels with Adobe Photoshop (2021), before 22 sections were placed into 20 Adobe Illustrator (2021) files for mapping. Only one side of the brain (right side of the section) has been labelled to leave the other side unobscured. Regions of the brain were labelled using terminology adapted from that used in the Paxinos and Watson system of brain atlases. The section number is given in the top right-hand corner of the plate. A small line-diagram finder image in each atlas plate shows the position of the horizontal section (grey line) within the brain. Finished atlas plate files were exported to jpg for publishing.
The Brain of the Common Bottlenose Dolphin
The brain is shortened anteroposteriorly due to the compression of the cranial cavity to accommodate the sonar lens known as the melon. The brain is dominated by the highly folded (gyrified) cerebral cortex and the equally foliated cerebellar hemispheres. The two largest cranial nerves are the optic (2n) and vestibulocochlear (8n)(plates 15 and 16).
The Dolphin Cerebral Cortex
The cerebral cortex of the bottlenose dolphin is highly folded (gyrified). This makes gyral and sulcal nomenclature potentially complex and difficult, but it was possible to identify distinct anteroposteriorly running sulci at the superior part of the cortex (superior medial sulcus – sms, superior intermediate sulcus – sis, superior lateral sulcus – sls; plates 2, 3). These can be used to demarcate frontal and occipital gyri. On the medial surface of the brain there are clearly demarcated pericallosal and cingulate sulci (pcs, cgs, respectively; plate 8), while the lateral surface of the brain has a deep lateral or Sylvian fissure (lf) leading to a recessed insula cortex encircled by a periinsular sulcus (pis, plate 9), leading inferiorly to the limen insulae (plates 13, 14).
The isocortex is difficult to subdivide on cytoarchitectonic grounds alone because most of the isocortex looks alike, but there is sufficient MRI evidence (Berns et al., 2015) to identify presumptive visual cortex in the superior occipital lobe (Vis) and an auditory cortex (Au) on the superior bank of the temporal lobe, where transverse temporal gyri (TTG; plates 12, 13) can be identified. The presumptive somatosensory cortex (Som) should be on the anterior parietal lobe. Piriform (Pir - primary olfactory; plates 13 to 15) cortex is clearly identifiable at the posterolateral base of the frontal cortex near the limen insulae.
Nomenclature for periallocortical and allocortical regions which have obvious topographical relationships and/or cytoarchitectural similarities to cortical regions identified in rodent, carnivore and primate cerebral cortex (e.g. indusium griseum, orbital, cingulate, entorhinal, agranular insula, granular insula) has been adopted to reflect those similarities.
The hippocampal formation is small for the size of the brain, with a minute dentate gyrus (plate 10) and a structurally simple cornu Ammonis (plates 9 to 13). Nevertheless, all components of the hippocampal formation are present.
The corpus callosum (plate 9) and anterior commissure (plate 14) are also small, suggesting minimal interhemispheric transfer of information in this species.
The Dolphin Auditory System
As might be expected given the behavioural importance of audition and sonar in delphinids, the components of the auditory pathway are very large. The vestibulocochlear nerve (8n) is as large as the optic nerve (2n)(see plates 15 and 16 for comparison). Most notable of the brainstem auditory nuclei are: i) the paired cochlear nuclear complexes (each comprising dorsal cochlear - DC, ventral cochlear anterior - VCA, and ventral cochlear posterior – VCP nuclei; see plates 15, 16); ii) the superior olivary nuclear complexes (medial superior olivary nucleus – MSO, lateral superior olivary nucleus - LSO, superior periolivary nucleus – SPO; see plates 15, 16); and iii) the inferior colliculi (plates 11 to 13).
The superior olivary nuclear complex probably plays a key role in spatial location of sound sources by comparing input from the two ears and by analysing echo delay. Components of the large inferior colliculi are structurally similar to other mammals, with each consisting of a core nucleus (CIC) surrounded by cortical regions (DCIC, ECIC). The inferior colliculus is probably key in using data from the superior olivary complex to map the position of sound sources in three-dimensional space. The outflow tracts from both the superior olive and inferior colliculus are very large (lateral lemniscus and brachium of inferior colliculus, respectively).
The auditory nucleus of the thalamus (medial geniculate nucleus; components MGD, MGV, MGMZ; see plates 11, 12) is also large, but not as striking as the superior olive and inferior colliculus, and the gyri of the presumptive auditory cortex on the temporal lobe (transverse temporal gyri – TTG; plates 12,13) don’t look much different from other mammals.
The overall impression is that much of the behaviourally important auditory processing in this species occurs at brainstem level (i.e. superior olive and inferior colliculus).
Cerebellar Systems in the Dolphin Brain
The cerebellar vermis and flocculus and their attendant deep cerebellar nuclei are structurally much like other mammals, suggesting that vestibulo- and spinocerebellar systems are similar to artiodactyls, primates and even rodents.
By contrast the corticopontocerebellar system is enlarged, with large cerebellar hemispheres, large lateral deep cerebellar nuclei (plates 13 to 15), and pontine nuclei extending from rhombomere 2 to 5 (plates 15 to 18). The dentatorubral and dentatothalamic tracts (plate 14) arising from the lateral deep cerebellar nucleus and forming the superior cerebellar peduncle, are also enlarged.
It is unlikely that the delphinid corticopontocerebellar system is serving the same appendicular motor functions as in primates, so it is possible that the expansion of this system is involved in some way with processing of sonar information and the behavioural demands of pelagic hunting.
Acknowledgements
I would like to thank Elizabeth Lockett of the National Museum of Health and Medicine in Silver Springs, Maryland, USA for the opportunity to study the marine mammal brains in the NMHM collection and for use of the Epson 11000scanner.
Reference
Berns GS, Cook PF, Foxley S, Jbabdi S, Miller KL, Marino L (2015) Diffusion tensor imaging of dolphin brains reveals direct auditory pathway to temporal lobe. Proceedings of the Royal Society, London. Series B. Issue 1811 Volume 282, DOI: 10.1098/rspb.2015.1203